
ЛЕКЦИЯ № 9 Липиды. Метаболизм Екатеринбург, 2016 г.


 - называются карбоновые кислоты, которые образуются при гидролизе")


 используются для синтеза БАВ")







 Fe 2+ (b 5) O 2 -")
 количественно")
")






, гипоксии, ишемии (при")
























 — стероид, характерный только для животных организмов. В сутки в")

 Биосинтез (эндогенный) Синтез липопротеидов Катаболизм Биомембраны Холекальциферол Стероидные гормоны")



")

, которые более гидрофобны, чем сам")



biohimiya._lekciya_9._lipidy._metabolizm._2016.ppt
- Размер: 810.5 Кб
- Автор:
- Количество слайдов: 60
Описание презентации ЛЕКЦИЯ № 9 Липиды. Метаболизм Екатеринбург, 2016 г. по слайдам
ЛЕКЦИЯ № 9 Липиды. Метаболизм Екатеринбург, 2016 г. Дисциплина: Биохимия Лектор: Гаврилов И. В. Факультет: лечебно-профилактический, Курс: 2 ФГБОУ ВО УГМУ Минздрава России Кафедра биохимии
Жирные кислоты • Жирными кислотами (ЖК) — называются карбоновые кислоты, которые образуются при гидролизе омыляемых липидов. • В основном к жирным кислотам относятся высшие карбоновые кислоты (содержащие 12 и более атомов С). • Высшие ЖК водонерастворимыми, они транспортируются в крови с помощью альбуминов, а в клетках — с помощью Z -белков.
№ Жирная кислота Индекс ЖК ∆ ЖК ω ЖК 1 Лауриновая 12: 0 2 Миристиновая 14: 0 3 Пальмитиновая 16: 0 4 Пальмитолеиновая 16: 1 ∆ 9 ω9 5 Стеариновая 18: 0 6 Олеиновая 18: 1 ∆ 9 ω9 7 Линолевая 18: 2 ∆ 9, 12 ω6 8 Линоленовая 18: 3 ∆ 9, 12, 15 ω3 9 Октадекатетраеновая 18: 4 ∆ 5, 8, 11, 14 ω3 10 Арахиновая 20: 0 11 Гадолеиновая 20: 1 ∆ 9 ω9 12 Эйкозатриеновая 20: 3 ∆ 8, 11, 14 ω
№ Жирная кислота Индекс ЖК ∆ ЖК ω ЖК 13 Арахидоновая 20: 4 ∆ 5, 8, 11, 14 ω6 14 Эйкозапентаеновая 20: 5 ∆ 5, 8, 11, 14, 17 ω3 15 Бегеновая 22: 0 16 Эруковая 22: 1 ∆ 13 ω9 17 Андреновая 22: 4 ∆ 9, 12, 15, 18 ω6 18 Докозапентаеновая 22: 5 ∆ 4, 7, 10, 13, 16 ω6 19 Докозагексаеновая 22: 6 ∆ 4, 7, 10, 13, 16, 19 ω3 20 Лигноцериновая 24: 0 21 Невроновая 24: 1 ∆ 15 ω9 22 Цереброновая 24: 0 α-гидрокси ЖК
Биологическое значение ЖК 1. полиеновые ЖК (арахидоновая, эйкозапентаеновая, эйкозатриеновая) используются для синтеза БАВ – эйкозаноидов (простагландинов, простациклинов, тромбоксанов, лейкотриенов, липоксинов). 2. ЖК окисляются в аэробных условиях с образованием АТФ; 3. ЖК являются структурным компонентом омыляемых липидов: восков, глицеролипидов, сфинголипидов, эфиров холестерина.
АНАБОЛИЗМ ЖИРНЫХ КИСЛОТ • Источником ЖК в организме являются синтетические процессы, омыляемые липиды и пища. • ЖК, которые синтезируются в организме, называются заменимыми. Значительная их часть образуется в печени , в, меньшей степени — в жировой ткани и лактирующей молочной железе. • ЖК, которые не синтезируются в организме, но необходимы для него называются незаменимыми. Единственным источником незаменимых ЖК является пища. Они образуют понятие витамин F (линолевая, линоленовая и арахидоновая).
Метаболизм ЖК СООН ЖКПища ТГ, ФЛ, ЭХС Ткани. Углеводы Аминокислоты Ацетил-Ко. А АТФ, СО 2 , Н 2 О ТГ, ФЛ, ЭХС Липогенез Липолиз Гидролиз β -окисление Пальмитат синтетаза и др. ЦТК, ЦОФ Диеновые конъюгаты Гидроперекиси Эндоперекиси Малоновый диальдегид и др. ПОЛ СООН ЖКα -окисление СО
Анаболизм ЖК СООН Пальмитат С 16 Пальмитат синтетаза СООН Миристиновая С 14 СООН Лауриновая С 12β-окисление β -окисление СООН Стеарат С 18 СООН Арахинат С 20 Элонгаза СООН Пальмитолеат С 16 : 1 СООН Олеат С 18 : 1 СООН Гадолеинат С 20 : 1 Десатуразы. Углеводы Аминокислоты Жирные кислоты Ацетил-Ко. А β-окисление
Синтез пальмитата Глюкоза Ацетил-Ко. А ПВК НАДФН 2 Г л и к о л и з. П Ф Ш Малонил-Ко. А Пальмитат- синтетазный комплекс Пальмитиновая кислота
ãëþêîçà ãë-6ô ôð-6ô ÏÊ ÏÊ Àöåòèë-ÊîÀ ÙÓÊ + Àöåòèë-ÊîÀöèòðàòÙÓÊìàëàò ÍÀÄÔÍ2 ÍÀÄÔ +ÍÀÄÍ + ÍÀÄÍ 2 ìèòîõîíäðèÿ ìàëàò ÄÃìàëèê-ôåðìåíò öèòðàò ñèíòàçà òðàíñëîêàçà öèòðàòëèàçà ÑÎ 2 HS-ÊîÀ ÀÒÔÀÄÔ+Ôí Ìàëîíèë-ÊîÀ ÀÄÔ+Ôí ÀÒÔÑÎ 2 Àöåòèë-ÊîÀ êàðáîêñèëàçà ïàëüìèòàò ÍÀÄÔ+ ïàëüìèòàò ñèíòàçà ÍÀÄÔ + ÍÀÄÔÍ 2 ÏÔØ öèòîïëàçìà ÍÀÄÔÍ 2 èçîöèòðàò Äà à-Êà ÀÒÔ, ÍÀÄÍ 2 Ñóêöèíèë-ÊîÀ à-Êà Äà ÀÒÔ, ÍÀÄÍ 2 èçîöèòðàò à-Êà ÍÀÄÔ + ÍÀÄÔÍ 2 èçîöèòðàò Äà ÖÒÊ öèòðàò, èíñóëèí ïàëüìèòîèë-ÊîÀ, àäðåíàëèí, ãëþêàãîí èíä. èíñóëèí
Ацетил-Ко. А карбоксилаза Ацетил-Ко. А Малонил-Ко. ААТФ АДФ + Фн СО 2 + Цитрат, инсулин — Адреналин, глюкагон, — пальмитоил-Ко. А биотин Регуляторная реакция синтеза пальмитата 1. СО 2 + биотин + АТФ → биотин-СООН + АДФ + Фн 2. ацетил-Ко. А + биотин-СООН → малонил-Ко. А + биотин 2 стадии:
Пальмитатсинтетазный комплекс
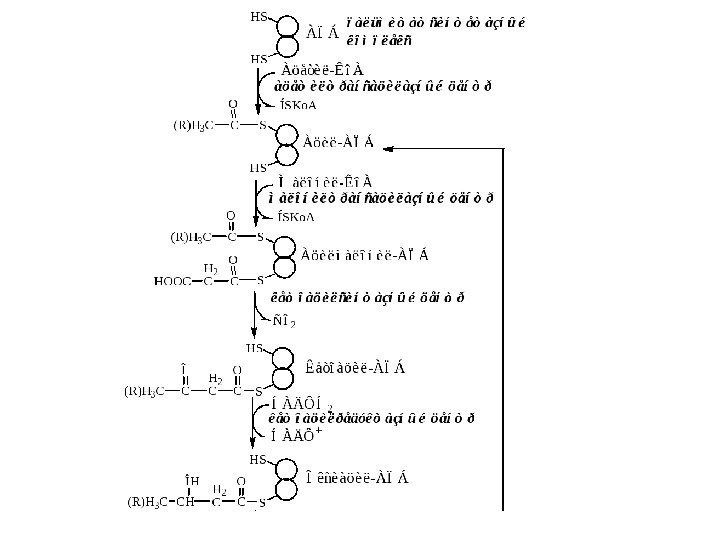
ДесатуразыÍÀÄÍ2 ÍÀÄ+ ÔÀÄÍ2 Fe 3+ (b 5) Fe 2+ (b 5) O 2 — + 2 H+H 2 O O 2 O Fe 3+ Fe 2+ ÄåñàòóðàçàÐåäóêòàçàÖèòîõðîìR H 2 CC RCH H 2 C OH R H CH 2 O SKo. A CSKo. A O O O Элонгазы 1. Удлинение ЖК называется элонгацией. 2. Для каждой длины ЖК существуют свои элонгазы (16 → 18, 18 → 20, 20 → 22, 22 → 24). Последовательность реакций аналогична синтезу пальмитиновой кислоты, однако в данном случае синтез идет не на АПБ, а на Ко. А. 3. Основной продукт элонгации в печени — стеариновая кислота. 4. В нервных тканях образуются ЖК с длинной цепью (С=20 -24), необходимые для синтеза сфинголипидов
Катаболизм ЖК • Хотя побочные пути (α-, ω-окисление ЖК, деградация ЖК в пероксисомах) количественно менее важны, их нарушение может приводить к тяжелым заболеваниям. Ферментативный Неферментативный • β-окисление (основной путь). • α-окисление • ω-окисление ЖК , • деградация ЖК в пероксисомах Перекисное окисление липидов (ПОЛ)
• ЖК с короткой и средней цепью (от 4 до 12 атомов С) могут проникать в матрикс митохондрий путём диффузии. Активация этих ЖК происходит ацил-Ко. А синтетазами в матриксе митохондрий. • ЖК с длинной цепью, сначала активируются в цитозоле (ацил-Ко. А синтетазами на внешней мембране митохондрий), а затем переносятся в матрикс митохондрий специальной транспортной системой с помощью карнитина. Транспорт ЖК в митохондрии L -карнитин • Карнитин поступает с пищей или синтезируется из лизина и метионина с участием витамина С.
Транспорт ЖК в митохондрии à ö è ë — Ê î À — ñ è í ò å ò à ç àH S — K o A R C OO S K o AR C O HO À Ò Ô À Ì Ô + Ô Ô íÆ è ð í à ÿ ê è ñ ë î ò à À ö è ë — S K o Aàöèë-ÊîÀ-ñèíòåòàçà HS-Ko. A RCO O SKo. ARCOH O ÀÒÔ ÀÌÔ+ÔÔí Æèðíàÿ êèñëîòàÀöèë-SKo. A Карнитин Н S- Ко. А Матрикс митохондрииβ -окисление Карнитинацил- трансфераза 1 Карнитинацил- трансфераза 2 Цитоплазма «-» Малонил-Ко. А
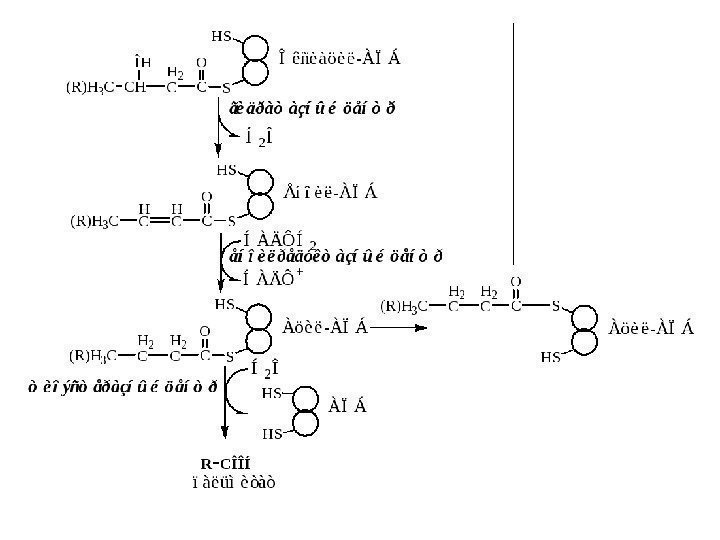
H 2 C CO S K o AH 2 CR À ö è ë — Ê î À H C CO S K o AH CR Å í î è ë — Ê î À H 2 C CO S K o AC ÍR Î ê ñ è à ö è ë — Ê î ÀÎ Í H 2 C CO S K o ACR Ê å ò î à ö è ë — Ê î ÀÎ Ô À Ä Í 2À ö è ë — Ê î À Ä Ã Í 2 Î Å í î è ë — Ê î À ã è ä ð à ò à ç à Î ê ñ è à ö è ë — Ê î À Ä ÃÍ À Ä + Í À Ä Í 2 CO S K o AR À ö è ë — Ê î À H 3 C CO S K o AÍ S K o A Ê å ò î à ö è ë — Ê î À ò è î ë à ç à Ö Ò Ê 8 À ö å ò è ë — Ê î ÀC = 1 6 , 1 4 , 1 2 , 1 0 , 8 , 6 , 4 C = 1 4 , 1 2 , 1 0 , 8 , 6 , 47 ö è ê ë î â C = 2 Ö Ï Ý 9 6 À Ò ÔÖ Ï Ý 7 Í À Ä Í 2 Ö Ï Ý 2 1 À Ò Ô 1 4 À Ò Ô 7 Ô À Ä Í 2 1 3 1 À Ò Ôï à ë ü ì è ò è ë — Ê î À C = 1 6 Реакции β-окисление ЖК
CH 3 H 2 C CO SKo. A COOH HC CO SKo. A CH 3 COOH CH CO SKo. A H 3 C COOH H 2 C CO SKo. A CH 2 C O 2 À Ä Ô + Ô íÀ Ò Ô Ï ð î ï è î í è ë — Ê î À D — Ì å ò è ë ì à ë î í è ë — Ê î À L — Ì å ò è ë ì à ë î í è ë — Ê î À Ñ ó ê ö è í è ë — Ê î À Ö Ò Ê Ö Ï Ý 6 À Ò ÔÏ ð î ï è î í è ë — Ê î À — ê à ð á î ê ñ è ë à ç àá è î ò è í Ì å ò è ë ì à ë î í è ë — Ê î À — ð à ö å ì à ç à Ì å ò è ë ì à ë î í è ë — Ê î À — è ç î ì å ð à ç à 12β-окисление насыщенной ЖК с нечетным количеством атомов С Последний цикл β-окисления сопровождается образованием не Ацетил-Ко. А, а Пропионил-Ко. А с 3 атомами С. Повреждение НСМетилмалоновая кислота
Регуляция β-окисление активируют : глюкагон, адреналин, ЖК, НАД+, АДФ: Голод, физическая нагрузка → ↑ глюкагон, ↑ адреналин → липолиз ТГ в адипоцитах → ↑ ЖК в крови → ↑ β-окисление в аэробных условиях в мышцах, печени → ↑АТФ; ↑ АТФ, ↑НАДH 2 ↑ Ацетил-Ко. А → ↑ цитрат, ↑ЖК → ↓ гликолиз → ↑ экономию глюкозы для нервной ткани, эритроцитов и т. д. β-окисление ингибируют: инсулин, НАДH 2 , АТФ. Пища → ↑ инсулин → ↑ гликолиз → ↑ Ацетил-Ко. А → ↑ синтез малонил-Ко. А и ЖК, ↑ малонил-Ко. А → ↓ карнитинацилтрансферазы I в печени → ↓ транспорт ЖК в матрикс митохондрий → ↓ ЖК в матриксе → ↓ β-окисление ЖК
C H 2 Ñ Î Î Í Ñ Ñ C H 2 Ñ Î Î Í Ï î ë è í å í à ñ û ù å í í à ÿ æ è ð í à ÿ ê è ñ ë î ò à Ý í ä î ï å ð å ê è ñ ü æ è ð í î é ê è ñ ë î ò û* O — O * O O C H H C C H 2 Ñ H H Ñ C H 2 Ñ Î Î Í Ì à ë î í î â û é ä è à ë ü ä å ã è äO O O OПОЛ объединяет все реакции неферментативного окисления полиненасыщенных ЖК, свободных или входящих в состав омыляемых липидов, протекающих по радикальному механизму. Реакции ПОЛ инициируются активными формами кислорода.
C H 2 Ñ Î Î Í H 2 O H * C C O O HC H 2 C H * Ñ Î Î ÍH Î * Ï î ë è í å í à ñ û ù å í í à ÿ æ è ð í à ÿ ê è ñ ë î ò à Ð à ä è ê à ë ï î ë è í å í à ñ û ù å í í î é æ è ð í î é ê è ñ ë î ò û Ä è å í î â û å ê î í ü þ ã à ò û æ è ð í û õ ê è ñ ë î òC H 2 H C Ñ Î Î Í O O * à è ä ð î ï å ð å ê è ñ ü æ è ð í î é ê è ñ ë î ò û Î2è í è ö è è ð î â à í è å 1 0 -6 ñ å ê ï å ð î ê ñ è ä í û é ð à ä è ê à ë æ è ð í î é ê è ñ ë î ò û C H 2 H C Ñ Î Î Í O O H R — C O O H R * — C O O HÎ ê è ñ ë å í è å í î â î é Æ Ê è ç î ì å ð è ç à ö è ÿ C H 2 H C Ñ Î Î Í O *H O * ñ ï î í ò à í í û é ð à ñ ï à ä F e 2+È í ä ó ö è ð î â à í í û é ð à ñ ï à ä F e 3+ O H — à è ä ð î ê ñ è ë ü í û é ð à ä è ê à ë æ è ð í î é ê è ñ ë î ò û C H 2 H C Ñ Î Î Í O H R — C O O H R* — C O O HÎ ê è ñ ë å í è å í î â î é Æ Ê ã è ä ð î ê ñ è æ è ð í à ÿ ê è ñ ë î ò àÐ Å À Ê Ö È È Ï Î Ë
Регуляция ПОЛ Процессы ПОЛ усиливаются при избытке катехоламинов (стресс), гипоксии, ишемии (при реоксигенации), повышенном содержании активных форм О 2 , снижении антиоксидантной защиты, повышенном содержании ненасыщенных жирных кислот. Активация ПОЛ происходит при воспалении и характерна для многих заболеваний: дистрофии мышц (болезнь Дюшенна), болезни Паркинсона, атеросклерозе, развитии опухолей. Физиологическое значение реакций ПОЛ: 1. модифицируют физико-химические свойства биомембран: увеличивают их проницаемость. 2. регулируют активность мембранных ферментов, реакции окислительного фосфорилирования. 3. участвуют в контроле клеточного деления. 4. Участвует в адаптации организма. Повышение активности ПОЛ приводит к: 1. разрушению, фрагментации клеточных мембран, повреждению и гибели клеток. 2. модификации ЛП, особенно ЛПНП. Они становятся «липкими» , легче проникают в сосудистую стенку, хорошо захватываются макрофагами, что ускоряет развитие атеросклероза. 3. накоплению продуктов ПОЛ, многие из которых токсичны, канцерогенены и мутагенены (МДА). 4. ускорению процессов старения организма.
Триглицериды Остатки кислот, % по массе Пальмитинов ая Стеаринов ая Олеиновая Линолевая Линол еновая Сливочное масло 25 11 34 6 5 Подсолнечное масло 11 4 38 46 — Оливковое масло 10 2 82 4 — Льняное масло 5 3 5 62 25 Бараний жир (твёрдый) 38 30 35 3 9 Говяжий жир (твёрдый) 31 26 40 2 2 Свиной жир (твёрдый) 27 14 45 5 5 ТГ в организме человека 25 8 46 10 —
Жировая ткань • Есть везде. Основные жировые отложения имеются под кожей (в подкожножировой клетчатке), и вокруг внутренних органов, преимущественно в области живота (висцеральный жир). Жировая ткань – это разновидность соединительной ткани, составляет 15 -20% веса мужчин и 20 -25% веса женщин. Как и любая ткань, она состоит из клеток (адипоциты, макрофаги, тучные клетки и т. д. ) и межклеточного вещества. Межклеточное вещество в жировой ткани представлено небольшим количеством волокон (коллагена и эластина) и основного вещества (гликозаминогликаны, протеоликаны). Белая жировая ткань
функции • Синтез липидов из углеводов • запасающая (95% ТГ в жировой ткани, 5% — во внутренних органах и костях), накопление витаминов А, Д, Е • теплоизоляционная ; • механическая защита органов и тканей; • эндокринная (секреция: лептина, эстрогенов, ангиотензиногена, интерлейкина-6, ФНО-α); • Антибактериальная защита (комплементарные факторы: адипсин, комплемент С 3, фактор В) • Регуляция воспаления (α 1 -кислый гликопротеин, гаптоглобин) • пассивное обезвреживание (депонирование) токсичных веществ • Участие в водно-солевом обмене • Обеспечивает поддержание гомеостаза
Химический состав 65 -85% — ТГ, 22% — вода, 5, 8% белок, 15 ммоль/кг калий. Из жирных кислот 42— 51% — олеиновая (18: 1), 22— 31% пальмитиновая (16: 0), 5— 14% пальмитоолеиновая (16: 1), 3— 5 % миристиновая, 1— 5 % линолевая
ОСОБЕННОСТИ МЕТАБОЛИЗМА АДИПОЦИТА БЕЛОЙ ЖИРОВОЙ ТКАНИ • Энергетический обмен медленный, анаэробный, потребляет мало кислорода. Энергия АТФ тратится на транспорт жирных кислот через клеточные мембраны (с участием карнитина). • Белковый обмен низкий, белки синтезируются адипоцитами преимущественно для собственных нужд. На экспорт синтезируются лептин, белки острой фазы воспаления (α 1 -кислый гликопротеин, гаптоглобин), компоненты системы комплимента (адипсин, комплемент С 3, фактор В), интерлейкины. • Углеводный обмен. Невысокий, преобладает катаболизм. • Липидный обмен — интенсивно идут реакции липолиза и липогенеза.
Бурая жировая ткань • Мало у взрослого человека, много у новорожденного • Локализована около почек и щитовидной железы. • Смешанная жировая ткань: между лопатками, на грудной клетке и плечах.
Функция бурой жировой ткани • термогенез • «взрывной» липолиз
ОСОБЕННОСТИ МЕТАБОЛИЗМА БУРОЙ ЖИРОВОЙ ТКАНИ 1. потребляет много кислорода 2. активно окисляет глюкозу и жирные кислоты 3. энергетический обмен высокий (низкий синтез АТФ, 2 реакции в гликолизе, 1 в ц. Кребса), в основном энергия в виде тепла (несократительный термогенез) 4. разобщение в митохондриях белком термогенином (РБ-1) процессов окисления и фосфорилирования 5. Характерен феномен «взрывного липолиза» 6. Термогенез активируется при переохлаждении, излишке липидов в крови, под действием лептина.
Липогенез – синтез липидов
Ацилтрансфераза 2 -МГ + Ацил~Ко. А → 1, 2 -ДГ + HS-Ко. А, 1, 2 -ДГ + Ацил~Ко. А → ТГ + HS-Ко. А 1, 2 -ДГ + ЦДФ-холин → лецитин + ЦМФ 1, 2 -ДГ + ЦДФ-этаноламин → кефалин + ЦМФ В клетках слизистой оболочки тонкой кишки синтезируются в основном видоспецифичные ТГ. Однако при поступлении с пищей ТГ с необычными жирными кислотами, например бараньего жира, в адипоцитах появляются ТГ, содержащие кислоты, характерные для бараньего жира (насыщенные разветвлённые жирные кислоты). 1. Моноацилглицероловый путь синтеза ТГ и ФЛàöèë-ÊîÀ-ñèíòåòàçà HS-Ko. A RCO O SKo. ARCOH O ÀÒÔ ÀÌÔ+ÔÔí Æèðíàÿ êèñëîòàÀöèë-SKo. A Только в энтероцитах
глюкоза 2. Глицерофосфатный путь синтеза ТГ и ФЛ Основной путь синтеза липидов в организмеàöèë-ÊîÀ-ñèíòåòàçà HS-Ko. A RCO O SKo. ARCOH O ÀÒÔ ÀÌÔ+ÔÔí Æèðíàÿ êèñëîòàÀöèë-SKo. A CH 2 OH Ñ CH 2 OPO 3 H 2 OHH ÍÀÄÍ2 ÍÀÄ+ Ãëèöåðîë-ô Äà ÔÄÀãëèöåðî-ôгликолиз 3 Í2Î RCOH O Æèðíàÿ êèñëîòà O O O Ñ Ñ Ñ O R 1 R 2 R 3 O O H 2 C HC H 2 C Òà ËÏË OH OH OH H 2 C HC H 2 C ãëèöåðèí +3 кровь цитоплазма + инсулин Глицерокиназа Только в печени. Печень, адипоцит OH OH OH H H OH CH 2 OH H АТФ АДФ
CH 2 OH Ñ CH 2 OPO 3 H 2 OHH R-CO-SKo. A HS-Ko. A Àöèëòðàíñôåðàçà ãëèöåðî-ô CH 2 O Ñ CH 2 OPO 3 H 2 OHH ëèçîôîñôàòèä C O Rмитохондрии R — C O — S K o A H S — K o A À ö è ë ò ð à í ñ ô å ð à ç àC H 2 O Ñ C H 2 O P O 3 H 2 O HH ë è ç î ô î ñ ô à ò è äCO R C H 2 O Ñ C H 2 O P O 3 H 2 OH ô î ñ ô à ò è äCO R Í2ÎÍ3ÐÎ4 ôîñôîòèäàò ôîñôîãèäðîëàçà + CH 2 O Ñ CH 2 OPO 3 H 2 OH ôîñôàòèä C O R CH 2 O Ñ CH 2 OH OH 1, 2 -ÄÃ C O R
R-CO-SKo. A HS-Ko. A Àöèëòðàíñôåðàçà CH 2 O C CH 2 OH OH 1, 2 -ÄÃ C O R CH 2 O C CH 2 O OH ÒÃ C O RH 2 C C H H 2 CO O P OCR 2 C H 2 N ( C H 3 ) 3 O HO O O C R 1 O L –Фосфатидилэтаноламин ЦДФ-холин ЦМФ Холин- трансфераза H 2 C CH H 2 C O OPO CR 2 CH 2 NH 2 OH O O OCR 1 OЦМФ ЦДФ- этаноламин L –Фосфатидилхолин (Лецитин)
Липолиз – распад липидов
Липолиз ТГCH 2 O C CH 2 OH OH 1, 2 -Äà C O R CH 2 O C CH 2 O OH Òà C O R CH 2 OH C CH 2 OH OH 2 -Ìà C O R CH 2 OH C CH 2 OH OHH ãëèöåðèí Í2Î R-COOHÍ2Î R-COOH ÒÀÃ-ëèïàçàÄèãëèöåðîëëèïàçàÌîíîãëèöåðîëëèïàçà • Липолиз в жировой ткани активируется при дефиците глюкозы в крови (постабсорбционный период, голодание, физическая нагрузка). • Процесс стимулируется глюкагоном, адреналином, в меньшей степени СТГ и глюкокортикоидами. • В результате липолиза концентрация свободных жирных кислот в крови возрастает в 2 раза.
Липолиз ФЛCH 2 CH H 2 C O O OPO C C R 1 R 2 CH 2 N(CH 3)3 OH O O O ÔË À 1 ÔË À 2 ÔË CÔË D Ôîñôàòèäèëõîëèí ëèçîÔË Ca 2 + Холин Фосфорная кислота. Жирные кислоты Глицерин
Кетоновые тела β-оксибутират ацетоацетат ацетон
Биологическое значение КТ 1. КТ — топливные молекулы, окисление β-гидроксибутирата до СО 2 и Н 2 О обеспечивает синтез 26 АТФ. Окисление КТ, как и ЖК сберегает глюкозу, что имеет большое значение в энергоснабжении аэробных тканей при голодании и физических нагрузках, когда возникает дефицит глюкозы. 2. Для нервной ткани КТ имеют исключительное значение, так как в отличие от мышц и почек, нервная ткань практически не использует ЖК в качестве источника энергии (ЖК не проходят гематоэнцефалический барьер). 3. Преимущество КТ перед ЖК: 1) КТ водорастворимы, а ЖК – нет; 2) ЖК разобщают окислительное фосфорилирование и усиливают синтез ТГ, а КТ – нет.
Схема обмена кетоновых тел Жирные кислоты АТФС мочой, выдыхаемым воздухом β-оксибутират ацетоацетат ацетон Кровь Печень О 2 СО 2 Ацил-Ко. А ацетоацетат β-оксибутират Тканиацетоацетат β-оксибутират
Синтез кетоновых тел в печени HS — Ko. A Тиолаза+ Ацетил-Ко. А ГМГ-Ко. АHS — Ko. A ГМГ-Ко. А синтаза Ацетил-Ко. А ГМГ-Ко. А лиаза Ацетоацетат. Митохондрии
Ацетоацетат β -оксибутират ацетон. СО 2 кровь НАДН 2 НАД +β -оксибутират ДГ
Катаболизм кетоновых тел Ацетоацетатβ -оксибутират НАДН 2 НАД + β -оксибутират ДГ 3 АТФ Ацетоацетат Сукцинат. Сукцинил-Ко. А ЦТК Сукцинил-Ко. А-ацетоацетат-Ко. А-трансфераза АТФ АДФ + Фн
Тиолаза Ацетил-Ко. АHS — Ko. A 12 АТФ Преимущество КТ перед ЖК: 1) КТ водорастворимы, а ЖК – нет; 2) ЖК разобщают окислительное фосфорилирование и усиливают синтез ТГ, а КТ – нет О
Нормы Содержание КТ в крови: • 30 мг/л – норма; • 300 -500 мг/л – при голодании; • 3 -4 г/л – при сахарном диабете;
ХОЛЕСТЕРИН Холестерин (ХС) — стероид, характерный только для животных организмов. В сутки в организме синтезируется 1 г ХС. ХС синтезируется в печени (50%), тонком кишечнике (15 -20%), коже, коре надпочечников, половых железах. С пищей поступает в сутки 0, 3— 0, 5 г ХС. Биологическая роль ХС • входит в состав мембран клеток, увеличивает их электроизоляционные свойства, придает жесткость и прочность; • В мембране защищает полиненасыщенные ЖК от окисления; • из ХС синтезируются жёлчные кислоты, стероидные гормоны, витамин Д 3 • является компонентом желчи, участвует в переваривании липидов.
Холестерин Эфир холестерина
Метаболизм холестерина Холестерин. Пища (экзогенный) Биосинтез (эндогенный) Синтез липопротеидов Катаболизм Биомембраны Холекальциферол Стероидные гормоны Желчные кислоты
СИНТЕЗ ХС происходит в цитозоле и ЭПР клеток. Это один из самых длинных метаболических путей в организме человека (около 100 последовательных реакций). 3 этапа: I этап — образование мевалоната (мевалоновой кислоты). • Две молекулы ацетил-Ко. А конденсируются тиолазой с образованием ацетоацетил-Ко. А; • Гидроксиметилглутарил-Ко. А-синтаза (ГМГ-Ко. А) присоединяет третий ацетильный остаток к ацетоацетил-Ко. А с образованием ГМГ-Ко. А. • ГМГ-Ко. А-редуктаза восстанавливает ГМГ-Ко. А до мевалоната с использованием 2 молекул НАДФH 2.
II этап — образование сквалена • Мевалонат превращается в изопреноидную структуру — изопентенилпирофосфат (5 атомов С); • 2 изопентенилпирофосфата конденсируются в геранилпирофосфат (10 атомов С); • Присоединение изопентенилпирофосфата к геранилпирофосфату дает фарнезилпирофосфат (15 атомов С). • 2 фарнезилпирофосфата конденсируются в сквален (15 атомов С).
III этап — образование ХС • Сквален циклазой превращается в ланостерин, (4 цикла и 30 атомов С). • Далее происходит 20 последовательных реакций, превращающих ланостерин в ХС (27 атомов С).
Синтез холестерина (ЭПР гепатоцитов-80%, энтероцитов-10%)
6 изопентенилпирофосфат
Этерификация ХС ХС образует с ЖК сложные эфиры (ЭХС), которые более гидрофобны, чем сам ХС. В клетках эту реакцию катализирует АХАТ(Ацил. Ко. А: ацилхолестеринтрансферазой): ХС + Ацил-Ко. А → ЭХС + HSKo. A ЭХС формирует в цитоплазме липидные капли, которые являются формой хранения ХС. По мере необходимости ЭХС гидролизуются холестеролэстеразой на ХС и ЖК. ЭХС синтезируются в крови в ЛПВП под действием ЛХАТ (лецитин: холестеролацилтрансферазой): ХС + лецитин → ЭХС + лизолецитин
Регуляция ключевого фермента синтеза ХС Гидрокси. Метил. Глутарил-Ко. А-редуктазы • Инсулин через дефосфорилирование активирует фермент. • Повышение концентрации ацетил-Ко. А стимулирует синтез ХС. • Синтез ХС активируется при питании углеводами и снижается при голодании. • Глюкагон через фосфорилирование ингибирует фермент. • ХС, желчные кислоты (в печени) репрессируют ген ГМГ-Ко. А-редуктазы.
Выведение ХС из организма В сутки из организма выводится 1, 0 г — 1, 3 г ХС. С желчью (0, 5 — 0, 7 г/сут) — в виде жёлчных кислот. Часть ХС в кишечнике под действием ферментов бактерий восстанавливается по двойной связи, образуя холестанол и копростанол. С кожным салом в сутки выделяется 0, 1 г ХС.
Спасибо за внимание!