

Введение, структура популяции.ppt
- Количество слайдов: 76
 Лекция 1. Введение в генетику популяций и количественных признаков
Лекция 1. Введение в генетику популяций и количественных признаков
 1 в. Понятие о генетике популяций, её предмет и цель исследований Генетика популяций – наука, изучающая закономерности динамики генетических процессов, происходящих в природных и экспериментальных популяциях живых организмов. Предмет этой науки – явления наследственности на популяционном уровне.
1 в. Понятие о генетике популяций, её предмет и цель исследований Генетика популяций – наука, изучающая закономерности динамики генетических процессов, происходящих в природных и экспериментальных популяциях живых организмов. Предмет этой науки – явления наследственности на популяционном уровне.
 сформулировать набор законов, отображающих переход от набора") Цель теории генетики популяций (Р. Левонтин, 1974) сформулировать набор законов, отображающих переход от набора генотипов к серии возможных фенотипов, с учётом действия естественного отбора, а также законов, которые позволяли бы по набору фенотипов в полученной популяции охарактеризовать представленные в ней генотипы. Переход от фенотипа к генотипу описывается с помощью генетических и эпигенетических законов, функциональной биологии и биологии развития. Популяционная генетика оперирует наборами генотипов, а биометрическая (количественная) генетика – наборами фенотипов изучаемых объектов.
Цель теории генетики популяций (Р. Левонтин, 1974) сформулировать набор законов, отображающих переход от набора генотипов к серии возможных фенотипов, с учётом действия естественного отбора, а также законов, которые позволяли бы по набору фенотипов в полученной популяции охарактеризовать представленные в ней генотипы. Переход от фенотипа к генотипу описывается с помощью генетических и эпигенетических законов, функциональной биологии и биологии развития. Популяционная генетика оперирует наборами генотипов, а биометрическая (количественная) генетика – наборами фенотипов изучаемых объектов.
 2 в. Основные этапы развития науки, выдающиеся учёные в области генетики популяций и количественных признаков. 1 этап. Вторая половина 20 -х-конец 30 -х годов XX века. Происходило накопление данных о генетической гетерогенности популяций. Данный этап завершился выработкой представлений о полиморфизме популяций. 2 этап. 40 -е-середина 60 -х годов XX века. Происходило изучение механизмов поддержания генетического полиморфизма популяций. Появились и получили развитие представления о важной роли гетерозиса в формировании генетического полиморфизма.
2 в. Основные этапы развития науки, выдающиеся учёные в области генетики популяций и количественных признаков. 1 этап. Вторая половина 20 -х-конец 30 -х годов XX века. Происходило накопление данных о генетической гетерогенности популяций. Данный этап завершился выработкой представлений о полиморфизме популяций. 2 этап. 40 -е-середина 60 -х годов XX века. Происходило изучение механизмов поддержания генетического полиморфизма популяций. Появились и получили развитие представления о важной роли гетерозиса в формировании генетического полиморфизма.
 3 этап. Вторая половина 60 -х-конец 1970 -х годов XX века. Этот этап характеризуется широким применением белкового электрофореза для изучения полиморфизма популяций. 4 этап. С конца 1970 -х годов. Этот этап характеризуется широким применением ДНКтехнологий для изучения особенностей процессов происходящих в популяциях. Примерно с начала 1990 х годов стали использоваться специальные программы и вычислительная техника для анализа разнообразных типов генетических данных.
3 этап. Вторая половина 60 -х-конец 1970 -х годов XX века. Этот этап характеризуется широким применением белкового электрофореза для изучения полиморфизма популяций. 4 этап. С конца 1970 -х годов. Этот этап характеризуется широким применением ДНКтехнологий для изучения особенностей процессов происходящих в популяциях. Примерно с начала 1990 х годов стали использоваться специальные программы и вычислительная техника для анализа разнообразных типов генетических данных.
 С. С. Четвериков В. Л. Иоганнсен
С. С. Четвериков В. Л. Иоганнсен
 С. С. Четвериков выдвинул положение о насыщении видов в природе возникающими мутациями и подчеркнул значение генетических процессов (мутация, свободное скрещивание, естественный отбор) и изоляции в видообразовании и эволюции, тем самым связав теорию эволюции Дарвина и генетики. Заложил основы эволюционной генетики. В работах 20 -х гг. С. С. Четвериков обосновывает три основные посылки популяционной генетики: ü Мутационный процесс в природных условиях протекает точно так же, как и в условиях лаборатории. Поэтому можно распространять выводы, полученные в лаборатории, на природные ситуации. ü Новые мутации возникают непрерывно и являются рецессивными по отношению к аллелям дикого типа, распространенным в природных популяциях. ü Характерной чертой природных популяций является преобладание в них панмиксии, что делает возможным приложение закона Харди-Вайнберга. С. С. Четвериков вывел породу шелкопряда – «Горьковская моновольтинная» .
С. С. Четвериков выдвинул положение о насыщении видов в природе возникающими мутациями и подчеркнул значение генетических процессов (мутация, свободное скрещивание, естественный отбор) и изоляции в видообразовании и эволюции, тем самым связав теорию эволюции Дарвина и генетики. Заложил основы эволюционной генетики. В работах 20 -х гг. С. С. Четвериков обосновывает три основные посылки популяционной генетики: ü Мутационный процесс в природных условиях протекает точно так же, как и в условиях лаборатории. Поэтому можно распространять выводы, полученные в лаборатории, на природные ситуации. ü Новые мутации возникают непрерывно и являются рецессивными по отношению к аллелям дикого типа, распространенным в природных популяциях. ü Характерной чертой природных популяций является преобладание в них панмиксии, что делает возможным приложение закона Харди-Вайнберга. С. С. Четвериков вывел породу шелкопряда – «Горьковская моновольтинная» .
 В. Вайнберг Г. Харди
В. Вайнберг Г. Харди
 — немецкий врач, в 1908 году независимо от английского математика") Вильгельм Вайнберг (1862 -1937) — немецкий врач, в 1908 году независимо от английского математика Годфри Харди сформулировавший обобщение, известное как Закон Харди — Вайнберга. Основные труды В. Вайнберга касаются генетики человека, медицинской статистики и эпидемиологии, включая проблемы изучения близнецов, мутаций, и приложения законов наследования к популяциям.
Вильгельм Вайнберг (1862 -1937) — немецкий врач, в 1908 году независимо от английского математика Годфри Харди сформулировавший обобщение, известное как Закон Харди — Вайнберга. Основные труды В. Вайнберга касаются генетики человека, медицинской статистики и эпидемиологии, включая проблемы изучения близнецов, мутаций, и приложения законов наследования к популяциям.
 Л. Кавалли-Сфорца Э. Майр
Л. Кавалли-Сфорца Э. Майр
 — один из ведущих мировых специалистов по популяционной генетике. Основным") Луиджи Лука Кавалли-Сфорца (1922) — один из ведущих мировых специалистов по популяционной генетике. Основным его достижением является исследование генетического происхождения человека и развития разных рас и народов, результаты его трудов собраны в книгах «История и география генов человека» и «Гены, народы и языки» . Кавалли-Сфорца разбил человечество на десять основных генетических кластеров — восточные азиаты, европейцы, эскимосы, юго-восточные азиаты, коренное население Америки, жители тихоокеанских островов, южные азиаты и северные африканцы, суб-сахарские африканцы, народы койсанской группы и центральноафриканские пигмеи, аборигены Австралии. По его теории, современный человек произошёл из Африки, хотя до этого уже существовали другие группы людей (в частности неандертальцы Европы), которые не сохранились. В дальнейшем люди мигрировали на Ближний Восток, в Европу и Азию, и распространились далее по суше и побережью до Юго-восточной Азии и Америки, и на лодках до Австралии и островов. Применяя математические методы разложения генетического распределения на принципиальные компоненты, он выделил различные потоки миграции, от древних до поздних, и дал им интерпретацию, сопоставляя с археологическими и лингвистическими данными.
Луиджи Лука Кавалли-Сфорца (1922) — один из ведущих мировых специалистов по популяционной генетике. Основным его достижением является исследование генетического происхождения человека и развития разных рас и народов, результаты его трудов собраны в книгах «История и география генов человека» и «Гены, народы и языки» . Кавалли-Сфорца разбил человечество на десять основных генетических кластеров — восточные азиаты, европейцы, эскимосы, юго-восточные азиаты, коренное население Америки, жители тихоокеанских островов, южные азиаты и северные африканцы, суб-сахарские африканцы, народы койсанской группы и центральноафриканские пигмеи, аборигены Австралии. По его теории, современный человек произошёл из Африки, хотя до этого уже существовали другие группы людей (в частности неандертальцы Европы), которые не сохранились. В дальнейшем люди мигрировали на Ближний Восток, в Европу и Азию, и распространились далее по суше и побережью до Юго-восточной Азии и Америки, и на лодках до Австралии и островов. Применяя математические методы разложения генетического распределения на принципиальные компоненты, он выделил различные потоки миграции, от древних до поздних, и дал им интерпретацию, сопоставляя с археологическими и лингвистическими данными.
 — американский биолог. Майр разрабатывал проблемы систематики, прежде всего, концепцию") Эрнст Майр (1904 -2005) — американский биолог. Майр разрабатывал проблемы систематики, прежде всего, концепцию биологического вида. Работы Майра — в особенности «Систематика и происхождение видов» (1942) — оказали существенное влияние на развитие синтетической теории эволюции. Майр также исследовал механизмы видообразования. Ему принадлежит открытие механизма и создание теории перипатрического видообразования, суть которой в том, что новые виды формируются в малых популяциях, изолированных у самых границ географического ареала распространения древней, родительской популяции.
Эрнст Майр (1904 -2005) — американский биолог. Майр разрабатывал проблемы систематики, прежде всего, концепцию биологического вида. Работы Майра — в особенности «Систематика и происхождение видов» (1942) — оказали существенное влияние на развитие синтетической теории эволюции. Майр также исследовал механизмы видообразования. Ему принадлежит открытие механизма и создание теории перипатрического видообразования, суть которой в том, что новые виды формируются в малых популяциях, изолированных у самых границ географического ареала распространения древней, родительской популяции.
 Дж. Холдейн С. Райт
Дж. Холдейн С. Райт
 Дж. Б. С. Холдейн и Сьюалл Райт связали дарвиновскую теорию эволюции и учение Грегора Менделя о наследственности, основываясь на математических и статистических доказательствах, полученных из анализа количественных и качественных (уровня, размеров, воспроизведения и т. д. ) показателей мутаций. Д. Холдейн: üразработал математическую теорию моделирования гена и сцепления наследственных факторов, üисследовал количественную сторону естественного и искусственного отбора, благодаря чему показал, что элементарная единица эволюционного процесса — популяция, а не отдельная особь ( «Факторы эволюции» , 1932). üопределил частоту мутирования генов у человека (1935), üввёл в науке понятие «генетического груза» (1937). Основные исследования 1924 -1934 гг. были посвящены систематическому изучению темпа изменения популяции под влиянием отбора и мутаций.
Дж. Б. С. Холдейн и Сьюалл Райт связали дарвиновскую теорию эволюции и учение Грегора Менделя о наследственности, основываясь на математических и статистических доказательствах, полученных из анализа количественных и качественных (уровня, размеров, воспроизведения и т. д. ) показателей мутаций. Д. Холдейн: üразработал математическую теорию моделирования гена и сцепления наследственных факторов, üисследовал количественную сторону естественного и искусственного отбора, благодаря чему показал, что элементарная единица эволюционного процесса — популяция, а не отдельная особь ( «Факторы эволюции» , 1932). üопределил частоту мутирования генов у человека (1935), üввёл в науке понятие «генетического груза» (1937). Основные исследования 1924 -1934 гг. были посвящены систематическому изучению темпа изменения популяции под влиянием отбора и мутаций.
 М. Кимура К. Пирсон
М. Кимура К. Пирсон
 — японский биолог. Объединив теоретическую популяционную генетику с данными молекулярной") Мото Кимура (1924 -1994) — японский биолог. Объединив теоретическую популяционную генетику с данными молекулярной эволюции, он развил нейтральную теорию молекулярной эволюции (1968), в которой генетический дрейф выступает важнейшим фактором изменения частоты аллелей в популяции. Идея этой теории в том, что на молекулярном уровне большинство генетических изменений несут нейтральный характер по отношению к естественному отбору, что делало генетический дрейф главным фактором эволюции.
Мото Кимура (1924 -1994) — японский биолог. Объединив теоретическую популяционную генетику с данными молекулярной эволюции, он развил нейтральную теорию молекулярной эволюции (1968), в которой генетический дрейф выступает важнейшим фактором изменения частоты аллелей в популяции. Идея этой теории в том, что на молекулярном уровне большинство генетических изменений несут нейтральный характер по отношению к естественному отбору, что делало генетический дрейф главным фактором эволюции.
 Н. В. Тимофеев. Ресовский Ф. Г. Добжанский
Н. В. Тимофеев. Ресовский Ф. Г. Добжанский
 — русский биолог. Основные направления исследований: радиационная генетика, популяционная") Николай Владимирович Тимофеев-Ресовский (1900 -1981) — русский биолог. Основные направления исследований: радиационная генетика, популяционная генетика, проблемы микроэволюции. Одна из основных работ по генетике популяций: Тимофеев. Ресовский Н. В. , Яблоков А. В. , Глотов Н. В. Очерк учения о популяции. М. : Наука, 1973. 276 с. Феодосий Григорьевич Добжанский (1900 -1975) — русскоамериканский ученый-зоолог, генетик, энтомолог, эволюционист. üИсследовал размах и состав генной изменчивости в популяциях, в частности той ее части, которую составляют летальные и другие вредные мутации. üПредложил концепцию «балансовой» генетической структуры популяций согласно которой генетический полиморфизм, т. е. устойчивое сохранение в популяции 2 -х и более, генетически различных групп особей, а также высокая степень генетической изменчивости характерны для обычного состояния генетической структуры природных популяций и являются основой протекания эволюционного процесса.
Николай Владимирович Тимофеев-Ресовский (1900 -1981) — русский биолог. Основные направления исследований: радиационная генетика, популяционная генетика, проблемы микроэволюции. Одна из основных работ по генетике популяций: Тимофеев. Ресовский Н. В. , Яблоков А. В. , Глотов Н. В. Очерк учения о популяции. М. : Наука, 1973. 276 с. Феодосий Григорьевич Добжанский (1900 -1975) — русскоамериканский ученый-зоолог, генетик, энтомолог, эволюционист. üИсследовал размах и состав генной изменчивости в популяциях, в частности той ее части, которую составляют летальные и другие вредные мутации. üПредложил концепцию «балансовой» генетической структуры популяций согласно которой генетический полиморфизм, т. е. устойчивое сохранение в популяции 2 -х и более, генетически различных групп особей, а также высокая степень генетической изменчивости характерны для обычного состояния генетической структуры природных популяций и являются основой протекания эволюционного процесса.
 Н. П. Дубинин Р. Фишер
Н. П. Дубинин Р. Фишер
 — советский генетик. Областью научных интересов Н. П. Дубинина") Николай Петрович Дубинин (1906 -1998) — советский генетик. Областью научных интересов Н. П. Дубинина была общая и эволюционная генетика, а также применение генетики в сельском хозяйстве. Вместе с А. С. Серебровским показал дробимость гена, а также явление комплементарности гена. Опубликовал ряд важных научных работ по структуре и функциям хромосом, показал наличие в популяциях генетического груза — летальных и сублетальных мутаций. Также работал в области космической генетики, над проблемами радиационной генетики. Автор классического учебника по генетике для студентов биологических специальностей высших учебных заведений.
Николай Петрович Дубинин (1906 -1998) — советский генетик. Областью научных интересов Н. П. Дубинина была общая и эволюционная генетика, а также применение генетики в сельском хозяйстве. Вместе с А. С. Серебровским показал дробимость гена, а также явление комплементарности гена. Опубликовал ряд важных научных работ по структуре и функциям хромосом, показал наличие в популяциях генетического груза — летальных и сублетальных мутаций. Также работал в области космической генетики, над проблемами радиационной генетики. Автор классического учебника по генетике для студентов биологических специальностей высших учебных заведений.
 - английский генетик. Известен своими трудами в области математической") Сэр Рональд Фишер (1890 -1962) - английский генетик. Известен своими трудами в области математической статистики, евгеники, эволюционной генетики. Его первая книга «Генетическая теория и естественный отбор» (1930) посвящена синтезу дарвиновской теории отбора и генетики. Р. Фишер выдвинул концепцию прогрессивного отбора и инбридинга, попытался унифицировать учение об эволюции (теорема естественного отбора).
Сэр Рональд Фишер (1890 -1962) - английский генетик. Известен своими трудами в области математической статистики, евгеники, эволюционной генетики. Его первая книга «Генетическая теория и естественный отбор» (1930) посвящена синтезу дарвиновской теории отбора и генетики. Р. Фишер выдвинул концепцию прогрессивного отбора и инбридинга, попытался унифицировать учение об эволюции (теорема естественного отбора).
 3 в. Основные понятия в генетике популяций, генетическая структура популяций Популяция – это совокупность особей одного вида, заселяющих определённую территорию, свободно скрещивающихся друг с другом и в той или иной степени изолированных от других совокупностей особей данного вида. Вид – это основная систематическая единица, реально существующая в природе, занимающая определённый ареал и представляющая совокупность родственных по происхождению особей, качественно отличных от других видов и не скрещивающихся с ними.
3 в. Основные понятия в генетике популяций, генетическая структура популяций Популяция – это совокупность особей одного вида, заселяющих определённую территорию, свободно скрещивающихся друг с другом и в той или иной степени изолированных от других совокупностей особей данного вида. Вид – это основная систематическая единица, реально существующая в природе, занимающая определённый ареал и представляющая совокупность родственных по происхождению особей, качественно отличных от других видов и не скрещивающихся с ними.
 доминантного и рецессивного аллелей (А и а), частоты") Генетической структурой популяции называются соотношения (частоты) доминантного и рецессивного аллелей (А и а), частоты гомозиготных (АА, аа) и гетерозиготного Аа генотипов, определяемых в процентах или долях единицы.
Генетической структурой популяции называются соотношения (частоты) доминантного и рецессивного аллелей (А и а), частоты гомозиготных (АА, аа) и гетерозиготного Аа генотипов, определяемых в процентах или долях единицы.
 Равновероятная встречаемость гамет при свободном скрещивании в популяции называется панмиксией. Концентрация генов – это их относительная частота в популяции. Равновесием структуры панмиктической популяции называется сохранение в поколениях частот аллелей и генотипов.
Равновероятная встречаемость гамет при свободном скрещивании в популяции называется панмиксией. Концентрация генов – это их относительная частота в популяции. Равновесием структуры панмиктической популяции называется сохранение в поколениях частот аллелей и генотипов.
 Обозначение Расшифровка Частота доминантного аллеля А Частота рецессивного аллеля а Частота доминантного гомозиготного генотипа АА Частота гетерозиготного генотипа Аа частота рецессивного гомозиготного генотипа аа
Обозначение Расшифровка Частота доминантного аллеля А Частота рецессивного аллеля а Частота доминантного гомозиготного генотипа АА Частота гетерозиготного генотипа Аа частота рецессивного гомозиготного генотипа аа
 Обозначение Расшифровка Численность доминантного гомозиготного генотипа Численность гетерозиготного генотипа Численность рецессивного гомозиготного генотипа Общая численность
Обозначение Расшифровка Численность доминантного гомозиготного генотипа Численность гетерозиготного генотипа Численность рецессивного гомозиготного генотипа Общая численность
 Обозначения аллелей, генотипов, их частот и численностей при множественном аллелизме
Обозначения аллелей, генотипов, их частот и численностей при множественном аллелизме
 Лекция 2 Основные законы генетики популяций и их использование в селекции 1 в. Закон Харди-Вайнберга (1908 г): Если популяция находится в равновесии, то частоты генотипов и генов у родителей и потомства не меняются в ряду поколений. Формула закона Харди-Вайнберга:
Лекция 2 Основные законы генетики популяций и их использование в селекции 1 в. Закон Харди-Вайнберга (1908 г): Если популяция находится в равновесии, то частоты генотипов и генов у родителей и потомства не меняются в ряду поколений. Формула закона Харди-Вайнберга:
 Зависимость частот генотипов от частот аллелей в диплоидной популяции f 1 , f 2 , f 3 p
Зависимость частот генотипов от частот аллелей в диплоидной популяции f 1 , f 2 , f 3 p
 Следствия из закона Харди-Вайнберга: 1. Равновесный состав популяции зависит только от частоты аллеля А или а в исходной родительской популяции. 2. Доля гетерозигот в равновесной панмиктической популяции не может превосходить 0, 5. Этот максимум достигается при частотах аллелей в исходной родительской популяции p=q=0, 5. 3. Редкие аллели присутствуют в популяции главным образом в гетерозиготном, а не в гомозиготном состоянии.
Следствия из закона Харди-Вайнберга: 1. Равновесный состав популяции зависит только от частоты аллеля А или а в исходной родительской популяции. 2. Доля гетерозигот в равновесной панмиктической популяции не может превосходить 0, 5. Этот максимум достигается при частотах аллелей в исходной родительской популяции p=q=0, 5. 3. Редкие аллели присутствуют в популяции главным образом в гетерозиготном, а не в гомозиготном состоянии.
 4. Чем ниже частота аллеля, тем большая доля этого аллеля присутствует в популяции в гетерозиготном состоянии. - Частота рецесивного аллеля в гетерозиготах Пример Частота людей страдающих алькаптонурией равна 0, 000001, т. е. 1 на 1 млн. , следовательно: Т. о. число генов алькаптонурии в гетерозиготах примерно в 1000 раз больше, чем в гомозиготах.
4. Чем ниже частота аллеля, тем большая доля этого аллеля присутствует в популяции в гетерозиготном состоянии. - Частота рецесивного аллеля в гетерозиготах Пример Частота людей страдающих алькаптонурией равна 0, 000001, т. е. 1 на 1 млн. , следовательно: Т. о. число генов алькаптонурии в гетерозиготах примерно в 1000 раз больше, чем в гомозиготах.
 Алькаптонурия - наследственное заболевание, связанное с врожденным нарушением метаболизма ароматических аминокислот тирозина и фенилаланина, в результате чего в организме накапливается промежуточный продукт обмена этих аминокислот — гомогентизиновая кислота (алькаптон). При этом заболевании моча при выходе становится черной, после 30 лет образуется черный пигмент в в тканях хряща и кожи (охроноз), развивается артрит (позвоночника и других суставов) начиная с раннего периода взросления, возникают проблемы с сердцем, камни в почках, камни в простате. Это болезнь встречается редко, поражая 1 из 250000 - 1 млн. людей в мире. Больше распространена на определенных территориях Словакии (где регистрируется примерно 1 случай заболевания на 19000) и в Доминиканской республике. Алкаптонурия возникает вследствие мутации гена, кодирующего синтез оксидазы гомогентезиновой кислоты. Данная патология характеризуется аутосомно-рецессивным типом наследования. Алкаптонурией чаще болеют мужчины. Ген оксидазы гомогетинзиновой кислоты человека (HGD) локализован на длинном плече 3 хромосомы человека.
Алькаптонурия - наследственное заболевание, связанное с врожденным нарушением метаболизма ароматических аминокислот тирозина и фенилаланина, в результате чего в организме накапливается промежуточный продукт обмена этих аминокислот — гомогентизиновая кислота (алькаптон). При этом заболевании моча при выходе становится черной, после 30 лет образуется черный пигмент в в тканях хряща и кожи (охроноз), развивается артрит (позвоночника и других суставов) начиная с раннего периода взросления, возникают проблемы с сердцем, камни в почках, камни в простате. Это болезнь встречается редко, поражая 1 из 250000 - 1 млн. людей в мире. Больше распространена на определенных территориях Словакии (где регистрируется примерно 1 случай заболевания на 19000) и в Доминиканской республике. Алкаптонурия возникает вследствие мутации гена, кодирующего синтез оксидазы гомогентезиновой кислоты. Данная патология характеризуется аутосомно-рецессивным типом наследования. Алкаптонурией чаще болеют мужчины. Ген оксидазы гомогетинзиновой кислоты человека (HGD) локализован на длинном плече 3 хромосомы человека.
 Харди-Вайнберга Следует доказать два утверждения закона Харди. Вайнберга: 1. Доказать, что") Доказательство закона (теоремы) Харди-Вайнберга Следует доказать два утверждения закона Харди. Вайнберга: 1. Доказать, что частоты аллелей в родительской и дочерней популяции одинаковы. 2. Доказать, что частоты генотипов в родительской и дочерней популяции также одинаковы.
Доказательство закона (теоремы) Харди-Вайнберга Следует доказать два утверждения закона Харди. Вайнберга: 1. Доказать, что частоты аллелей в родительской и дочерней популяции одинаковы. 2. Доказать, что частоты генотипов в родительской и дочерней популяции также одинаковы.
 Результаты объединения мужских и женских гамет Мужские гаметы Женские гаметы p. A qa Сумма p q
Результаты объединения мужских и женских гамет Мужские гаметы Женские гаметы p. A qa Сумма p q
 Диаллельная схема равновероятного скрещивания особей трёх генотипов Материнские генотипы и их частоты Отцовские генотипы и их частоты
Диаллельная схема равновероятного скрещивания особей трёх генотипов Материнские генотипы и их частоты Отцовские генотипы и их частоты
 Частоты скрещиваний и частоты генотипов потомков по каждому из них Тип Частоты Потомство скрещивания АА Аа аа ния АА х АА 0 0 АА х Аа 0 Аа х Аа АА х аа 0 Аа х аа 0 аа х аа 0 Сумма 1, 00 0 0
Частоты скрещиваний и частоты генотипов потомков по каждому из них Тип Частоты Потомство скрещивания АА Аа аа ния АА х АА 0 0 АА х Аа 0 Аа х Аа АА х аа 0 Аа х аа 0 аа х аа 0 Сумма 1, 00 0 0
 Частота доминантного генотипа в дочерней популяции: Частота гетерозиготы: Частота рецессивного генотипа в дочерней популяции:
Частота доминантного генотипа в дочерней популяции: Частота гетерозиготы: Частота рецессивного генотипа в дочерней популяции:
 Условия выполнения закона Харди-Вайнберга: 1. Концентрация аллелей не должна изменяться под влиянием прямых и обратных мутаций, т. е. популяция не должна подвергаться давлению мутационного процесса. 2. 2. Популяция не должна испытывать на себе давления отбора. Особи, имеющие разные генотипы, должны обладать одинаковой жизнеспособностью и плодовитостью. 3. 3. На популяции не должно сказываться давление миграции, т. е. проникновение и вовлечение в скрещивание особей из других популяций с другим соотношением генов.
Условия выполнения закона Харди-Вайнберга: 1. Концентрация аллелей не должна изменяться под влиянием прямых и обратных мутаций, т. е. популяция не должна подвергаться давлению мутационного процесса. 2. 2. Популяция не должна испытывать на себе давления отбора. Особи, имеющие разные генотипы, должны обладать одинаковой жизнеспособностью и плодовитостью. 3. 3. На популяции не должно сказываться давление миграции, т. е. проникновение и вовлечение в скрещивание особей из других популяций с другим соотношением генов.
 4. Популяция должна иметь очень большую численность, чтобы на концентрации генов не сказались случайные отклонения, неизбежные при ограниченных выборках. 5. На популяцию не должны оказывать влияние факторы изоляции. Скрещивание между особями должно быть равновероятным.
4. Популяция должна иметь очень большую численность, чтобы на концентрации генов не сказались случайные отклонения, неизбежные при ограниченных выборках. 5. На популяцию не должны оказывать влияние факторы изоляции. Скрещивание между особями должно быть равновероятным.
 2 в. Закон Харди-Вайнберга для полиплоидов
2 в. Закон Харди-Вайнберга для полиплоидов
 значительная часть генотипов является гетерозиготами (симплексы,") В популяциях автотетраплоидов (сорта ржи, гречихи, кормовых трав) значительная часть генотипов является гетерозиготами (симплексы, дуплексы, триплексы). При p=q=0, 5 доля гомозигот (нулиплексы, квадриплексы) составляет незначительную часть популяции. У диплоидов же при p=q=0, 5 половина особей популяции – гомозиготы.
В популяциях автотетраплоидов (сорта ржи, гречихи, кормовых трав) значительная часть генотипов является гетерозиготами (симплексы, дуплексы, триплексы). При p=q=0, 5 доля гомозигот (нулиплексы, квадриплексы) составляет незначительную часть популяции. У диплоидов же при p=q=0, 5 половина особей популяции – гомозиготы.
 Равновесие у полиплоидов не может быть достигнуто за одно поколение размножения, поскольку у полиплоидов возрастает число генотипов гамет и для достижения равновесия требуется нескольких поколений размножения. Равновесные доли пяти генотипов в популяции автотетраплоидов задаются выражением:
Равновесие у полиплоидов не может быть достигнуто за одно поколение размножения, поскольку у полиплоидов возрастает число генотипов гамет и для достижения равновесия требуется нескольких поколений размножения. Равновесные доли пяти генотипов в популяции автотетраплоидов задаются выражением:
: 1. Если популяция не находится") 3 в. Закон стабилизирующего скрещивания К. Пирсона (1904 г): 1. Если популяция не находится в состоянии равновесия, то равновесие при случайном скрещивании достигается через одно поколение и не зависит от первоначального состава популяции. 2. Все популяции с одними и теми же частотами генов (хотя частоты генотипов могут и различаться) при случайном подборе пар придут к одному и тому же равновесному состоянию.
3 в. Закон стабилизирующего скрещивания К. Пирсона (1904 г): 1. Если популяция не находится в состоянии равновесия, то равновесие при случайном скрещивании достигается через одно поколение и не зависит от первоначального состава популяции. 2. Все популяции с одними и теми же частотами генов (хотя частоты генотипов могут и различаться) при случайном подборе пар придут к одному и тому же равновесному состоянию.
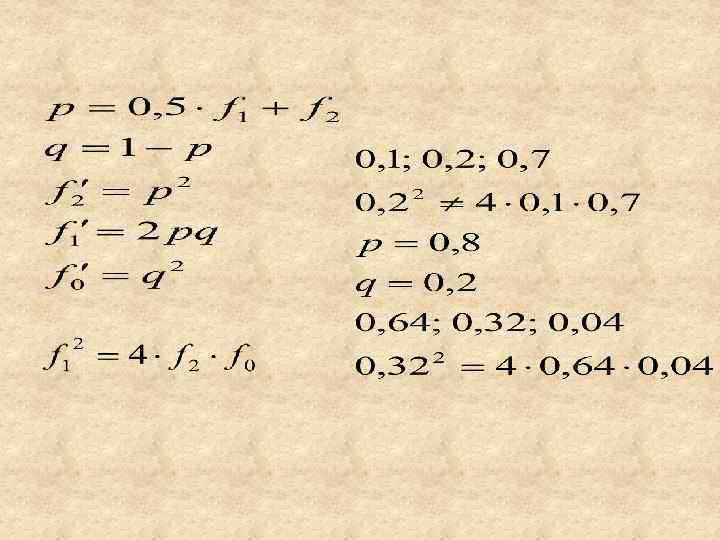
 4 в. Использование закономерностей генетической структуры популяций в селекционной практике. Закономерности генетической структуры популяций используются для решения следующих задач: 1. Определение частот генов по частотам или численностям генотипов в популяциях при неполном доминировании или кодоминировании. 2. Определение частот генов и генотипов по частоте или численности рецессивного генотипа при полном доминоровании. 3. Определение частоты гетерозигот как «носителей» какихлибо рецессивных признаков. 4. Проверка соответствия наблюдаемых частот генотипов теоретически ожидаемым по закону Харди-Вайнберга.
4 в. Использование закономерностей генетической структуры популяций в селекционной практике. Закономерности генетической структуры популяций используются для решения следующих задач: 1. Определение частот генов по частотам или численностям генотипов в популяциях при неполном доминировании или кодоминировании. 2. Определение частот генов и генотипов по частоте или численности рецессивного генотипа при полном доминоровании. 3. Определение частоты гетерозигот как «носителей» какихлибо рецессивных признаков. 4. Проверка соответствия наблюдаемых частот генотипов теоретически ожидаемым по закону Харди-Вайнберга.
 Для определения частот аллелей при неполном доминировании или кодоминировании, т. е. когда все генотипы можно распознать по фенотипу используют формулы максимального правдоподобия Р. Фишера:
Для определения частот аллелей при неполном доминировании или кодоминировании, т. е. когда все генотипы можно распознать по фенотипу используют формулы максимального правдоподобия Р. Фишера:
 Частоты генов и генотипов для двух аллелей одного локуса при полном доминировании определяют по формулам Харди-Вайнберга: Если в задаче дана численность рецессивного генотипа среди всех особей, то сначала нужно определить частоту рецессивного генотипа по формуле:
Частоты генов и генотипов для двух аллелей одного локуса при полном доминировании определяют по формулам Харди-Вайнберга: Если в задаче дана численность рецессивного генотипа среди всех особей, то сначала нужно определить частоту рецессивного генотипа по формуле:
 среди нормальных особей (доминантных гомозигот и гетерозигот)") Для определение частоты носителей рецессивных признаков (гетерозигот) среди нормальных особей (доминантных гомозигот и гетерозигот) используют следующую формулу:
Для определение частоты носителей рецессивных признаков (гетерозигот) среди нормальных особей (доминантных гомозигот и гетерозигот) используют следующую формулу:
 Для установления соответствия между фактическими частотами генотипов и теоретически ожидаемыми по закону Харди-Вайнберга проводят вычисления в следующем порядке: 1. Определяют частоту аллелей по формуле Р. Фишера. 2. 2. Рассчитывают теоретические частоты генотипов по формуле Харди-Вайнберга. 3. Определяют теоретические численности генотипов. 4. Рассчитывают критерий (хи-квадрат). 5. Находят табличное значение критерия хи-квадрат. Если расчетная величина хи-квадрат меньше табличной, то частоты генотипов в изучаемой популяции соответствуют равновесию Харди-Вайнберга.
Для установления соответствия между фактическими частотами генотипов и теоретически ожидаемыми по закону Харди-Вайнберга проводят вычисления в следующем порядке: 1. Определяют частоту аллелей по формуле Р. Фишера. 2. 2. Рассчитывают теоретические частоты генотипов по формуле Харди-Вайнберга. 3. Определяют теоретические численности генотипов. 4. Рассчитывают критерий (хи-квадрат). 5. Находят табличное значение критерия хи-квадрат. Если расчетная величина хи-квадрат меньше табличной, то частоты генотипов в изучаемой популяции соответствуют равновесию Харди-Вайнберга.
 рассчитывают по следующей формуле: где О и Е – наблюдаемые и") Критерий соответствия (хи-квадрат) рассчитывают по следующей формуле: где О и Е – наблюдаемые и теоретически ожидаемые количества генотипов определённого типа, а k – число генотипических (фенотипических) классов.
Критерий соответствия (хи-квадрат) рассчитывают по следующей формуле: где О и Е – наблюдаемые и теоретически ожидаемые количества генотипов определённого типа, а k – число генотипических (фенотипических) классов.
 Табличное значение хи-квадрат находят по числу степеней свободы и уровню значимости в специальной таблице. Степень свободы определяет значимость величины хиквадрат и равна числу фенотипических (генотипических) классов k минус единица и минус число оцениваемых параметров (число независимых величин). При кодоминировании число фенотипических классов, число оцениваемых параметров и число степеней свободы определяют по формулам:
Табличное значение хи-квадрат находят по числу степеней свободы и уровню значимости в специальной таблице. Степень свободы определяет значимость величины хиквадрат и равна числу фенотипических (генотипических) классов k минус единица и минус число оцениваемых параметров (число независимых величин). При кодоминировании число фенотипических классов, число оцениваемых параметров и число степеней свободы определяют по формулам:
 Уровень значимости обычно принимается равным 0, 05 д. е. или 5%. Это означает, что гипотезу решено считать не соответствующей наблюдениям, если вероятность того, что расхождение между теоретически ожидавшимися и наблюдаемыми в эксперименте данными, обусловленное только случайными причинами, составляет не более 5%.
Уровень значимости обычно принимается равным 0, 05 д. е. или 5%. Это означает, что гипотезу решено считать не соответствующей наблюдениям, если вероятность того, что расхождение между теоретически ожидавшимися и наблюдаемыми в эксперименте данными, обусловленное только случайными причинами, составляет не более 5%.
 Пример У людей имеются два аллеля в локусе. В выборке из 200 человек обладатели различных генотипов распределились следующим образом: - 108 - 86 -6 Всего 200 Следует установить, соответствуют ли эти данные частотам, которых следует ожидать, исходя из равновесия Харди-Вайнберга.
Пример У людей имеются два аллеля в локусе. В выборке из 200 человек обладатели различных генотипов распределились следующим образом: - 108 - 86 -6 Всего 200 Следует установить, соответствуют ли эти данные частотам, которых следует ожидать, исходя из равновесия Харди-Вайнберга.
 Частоты аллелей: Теоретические частоты генотипов: Теоретические численности генотипов:
Частоты аллелей: Теоретические частоты генотипов: Теоретические численности генотипов:
 Хи-квадрат фактический: Число степеней свободы: При числе степеней свободы df=1 и уровне значимости P=0, 05 (5%) хи-квадрат табличный ( ) равен 3, 84.
Хи-квадрат фактический: Число степеней свободы: При числе степеней свободы df=1 и уровне значимости P=0, 05 (5%) хи-квадрат табличный ( ) равен 3, 84.
, то величина 5,") Вывод: Поскольку фактический критерий хиквадрат больше табличного (5, 26 >3, 84), то величина 5, 26, статистически достоверна при 5%-ном уровне значимости. Следовательно следует отвергнуть гипотезу о соответствии частот указанных трёх генотипов равновесию Харди-Вайнберга.
Вывод: Поскольку фактический критерий хиквадрат больше табличного (5, 26 >3, 84), то величина 5, 26, статистически достоверна при 5%-ном уровне значимости. Следовательно следует отвергнуть гипотезу о соответствии частот указанных трёх генотипов равновесию Харди-Вайнберга.
 Лекция 3. Генетика раздельнополых популяций 1 в. Признаки, сцепленные с полом, гены гаплодиплоидов. Знание особенностей генетических процессов происходящих в раздельнополых популяциях важно для селекции двудомных раздельнополых растений и для медицины. Например, ряд опасных болезней человека сцеплены с Ххромосомой. К рецессивным признакам сцепленным с Х-хромосомой относятся: гемофилия, дальтонизм (неспособность различать красный и зелёный цвета), атрофия зрительного нерва и миопатия Дюшена. К доминантным признакам относят - рахит, не поддающийся лечению витамином Д, и тёмная эмаль зубов.
Лекция 3. Генетика раздельнополых популяций 1 в. Признаки, сцепленные с полом, гены гаплодиплоидов. Знание особенностей генетических процессов происходящих в раздельнополых популяциях важно для селекции двудомных раздельнополых растений и для медицины. Например, ряд опасных болезней человека сцеплены с Ххромосомой. К рецессивным признакам сцепленным с Х-хромосомой относятся: гемофилия, дальтонизм (неспособность различать красный и зелёный цвета), атрофия зрительного нерва и миопатия Дюшена. К доминантным признакам относят - рахит, не поддающийся лечению витамином Д, и тёмная эмаль зубов.
 самцы гаплоидны (гемизиготны) по аллелям Х -хромосомы, поэтому гетерозиготны") При гетерогаметности мужского пола (ХУ) самцы гаплоидны (гемизиготны) по аллелям Х -хромосомы, поэтому гетерозиготны по этим аллелям только самки (ХХ). Для гапло-диплоидов, например перепончатокрылых, где самцы гаплоидны, а самки диплоидны, гетерозиготность по всем локусам относится только к самкам.
При гетерогаметности мужского пола (ХУ) самцы гаплоидны (гемизиготны) по аллелям Х -хромосомы, поэтому гетерозиготны по этим аллелям только самки (ХХ). Для гапло-диплоидов, например перепончатокрылых, где самцы гаплоидны, а самки диплоидны, гетерозиготность по всем локусам относится только к самкам.
 2 в. Установление равновесного состояния в растительных популяциях при сцеплении с полом Система Х и У хромосом позволяет поддерживать равенство полов в популяции. При детерминации пола цветков у растений с Х и У хромосомами, растения с пестичными цветками, как правило гомогаметны (ХХ), а с тычиночными гетерогаметны (ХУ). В популяции раздельнополых растений (конопля, спаржа, шпинат и др. ) скрещиваются растения противоположных фенотипов (диссортативное скрещивание).
2 в. Установление равновесного состояния в растительных популяциях при сцеплении с полом Система Х и У хромосом позволяет поддерживать равенство полов в популяции. При детерминации пола цветков у растений с Х и У хромосомами, растения с пестичными цветками, как правило гомогаметны (ХХ), а с тычиночными гетерогаметны (ХУ). В популяции раздельнополых растений (конопля, спаржа, шпинат и др. ) скрещиваются растения противоположных фенотипов (диссортативное скрещивание).
 частоты аллелей и генотипов совпадают, если рассматриваемые гены") У растений с тычиночными цветками (ХУ) частоты аллелей и генотипов совпадают, если рассматриваемые гены локализованы в Х хромосоме (аллели генов находятся в гемизиготном состоянии): Для растений с пестичными цветками (ХХ) частоты генов и генотипов записываются как обычно (каждый ген в Х хромосоме имеет по два аллеля):
У растений с тычиночными цветками (ХУ) частоты аллелей и генотипов совпадают, если рассматриваемые гены локализованы в Х хромосоме (аллели генов находятся в гемизиготном состоянии): Для растений с пестичными цветками (ХХ) частоты генов и генотипов записываются как обычно (каждый ген в Х хромосоме имеет по два аллеля):
 При случайном переносе пыльцы и случайном оплодотворении частоты генов и генотипов находятся в равновесии, т. е. закон Харди. Вайнберга выполняется и для генов, сцепленных с половыми хромосомами. Однако, в случае сцепления с полом пестичные растения несут две дозы генов, а тычиночные – одну, поэтому равновесие частот генов и генотипов достигается только после нескольких поколений свободного размножения. Приближение к равновесию носит колебательный характер. Это связано с тем, что тычиночные растения получают Х хромосому от пестичного растения, а пестичные – от обоих родительских растений.
При случайном переносе пыльцы и случайном оплодотворении частоты генов и генотипов находятся в равновесии, т. е. закон Харди. Вайнберга выполняется и для генов, сцепленных с половыми хромосомами. Однако, в случае сцепления с полом пестичные растения несут две дозы генов, а тычиночные – одну, поэтому равновесие частот генов и генотипов достигается только после нескольких поколений свободного размножения. Приближение к равновесию носит колебательный характер. Это связано с тем, что тычиночные растения получают Х хромосому от пестичного растения, а пестичные – от обоих родительских растений.
 зависит от исходных частот в мужской ( ) и") Равновесная частота аллеля ( ) зависит от исходных частот в мужской ( ) и в женской ( ) частях популяции следующим образом: Пример: Популяция состоит из мужских и женских особей с контрастными частотами аллеля А:
Равновесная частота аллеля ( ) зависит от исходных частот в мужской ( ) и в женской ( ) частях популяции следующим образом: Пример: Популяция состоит из мужских и женских особей с контрастными частотами аллеля А:
 В результате первого свободного скрещивания А//А х а/ → А//а + А/ генетическая структура женской части популяции потомства определяется в равной степени частотами аллелей в обеих исходных частях:
В результате первого свободного скрещивания А//А х а/ → А//а + А/ генетическая структура женской части популяции потомства определяется в равной степени частотами аллелей в обеих исходных частях:
 После второго свободного скрещивания: и т. д.
После второго свободного скрещивания: и т. д.
 Поколения свободного скрещивания Исходное 1 2 3 4 5 6 7 8 1 0, 5 0, 75 0, 63 0, 69 0, 66 0, 67 0, 66
Поколения свободного скрещивания Исходное 1 2 3 4 5 6 7 8 1 0, 5 0, 75 0, 63 0, 69 0, 66 0, 67 0, 66
 Изменение частоты доминантного аллеля в ряду поколений свободного скрещивания в женской и мужской частях популяции при сцеплении с полом
Изменение частоты доминантного аллеля в ряду поколений свободного скрещивания в женской и мужской частях популяции при сцеплении с полом
 3 в. Установление равновесия популяции при межполовых различиях по частоте аллеля аутосомного локуса После первого свободного скрещивания: Мужские гаметы Женские гаметы
3 в. Установление равновесия популяции при межполовых различиях по частоте аллеля аутосомного локуса После первого свободного скрещивания: Мужские гаметы Женские гаметы
 Таким образом, частоты генотипов АА, Аа и аа после первого свободного скрещивания будут следующие: Равновесие популяции ещё не достигнуто, что видно из примера:
Таким образом, частоты генотипов АА, Аа и аа после первого свободного скрещивания будут следующие: Равновесие популяции ещё не достигнуто, что видно из примера:
 Равновесная частота доминантного аллеля равна: Однако эта частота не равна полученной при первом свободном скрещивании – 0, 18. Следовательно популяция ещё не находится в равновесном состоянии. Равновесие наступит только после ещё одного свободного скрещивания.
Равновесная частота доминантного аллеля равна: Однако эта частота не равна полученной при первом свободном скрещивании – 0, 18. Следовательно популяция ещё не находится в равновесном состоянии. Равновесие наступит только после ещё одного свободного скрещивания.
 4 в. Определение частот аллелей при сцеплении с полом При Х-сцепленном кодоминировании частота рецессивного аллеля определяется по следующим формулам: В случае Х-сцепленного доминирования частота рецессивного аллеля определяется по формулам:
4 в. Определение частот аллелей при сцеплении с полом При Х-сцепленном кодоминировании частота рецессивного аллеля определяется по следующим формулам: В случае Х-сцепленного доминирования частота рецессивного аллеля определяется по формулам:
 Пример для Х-сцепленного кодоминирования Рецессивный ген yellow - y, определяющий окраску шерсти у кошек сцеплен с Х-хромосомой. Гетерозиготные кошки (+y) имеют черепаховую окраску. Гомозиготные по аллелю y генотипы кошек (yy) и гемизиготные cамцы (y) имеют жёлтую окраску меха. Остальные кошки (++) и коты (+) имеют обычную окраску (дикий тип). Подсчёт животных в ветклинике дал следующие результаты: ++ - 277 +y – 54 yy– 7 + - 311 y - 42
Пример для Х-сцепленного кодоминирования Рецессивный ген yellow - y, определяющий окраску шерсти у кошек сцеплен с Х-хромосомой. Гетерозиготные кошки (+y) имеют черепаховую окраску. Гомозиготные по аллелю y генотипы кошек (yy) и гемизиготные cамцы (y) имеют жёлтую окраску меха. Остальные кошки (++) и коты (+) имеют обычную окраску (дикий тип). Подсчёт животных в ветклинике дал следующие результаты: ++ - 277 +y – 54 yy– 7 + - 311 y - 42
 Из полученных данных можно определить частоты рецессивного аллеля у кошек и котов:
Из полученных данных можно определить частоты рецессивного аллеля у кошек и котов:
 содержит сцепленный с полом") Пример для Х-сцепленного доминирования У людей локус ma (макроглобулин а) содержит сцепленный с полом ген, кодирующий макроглобулин сыворотки крови. Наличие в сыворотке крови этого антигена ( ) доминантно по отношению к его отсутствию ( ). В одной из выборок среди населения Норвегии распределение фенотипов было следующим: у женщин – 57 и 44 у мужчин – 23 и 77 Следовательно, частота аллеля у женщин:
Пример для Х-сцепленного доминирования У людей локус ma (макроглобулин а) содержит сцепленный с полом ген, кодирующий макроглобулин сыворотки крови. Наличие в сыворотке крови этого антигена ( ) доминантно по отношению к его отсутствию ( ). В одной из выборок среди населения Норвегии распределение фенотипов было следующим: у женщин – 57 и 44 у мужчин – 23 и 77 Следовательно, частота аллеля у женщин:
 Частота этого же аллеля у мужчин: Частота аллеля составляет: в целом по всей популяции
Частота этого же аллеля у мужчин: Частота аллеля составляет: в целом по всей популяции
 5 в. Определение частоты встречаемости рецессивных фенотипов в мужской части популяции по сравнению с женской частью популяции Поскольку частоты двух гемизиготных генотипов в мужской части популяции при сцеплении с полом совпадают с частотами соответствующих аллелей у женских особей предыдущего поколения, то фенотипы, определяемые рецессивными генами, у мужских особей будут встречаться чаще, чем у женских. Если частота сцепленного с полом рецессивного аллеля равна , то частота определяемого им фенотипа будет равна для мужских особей и для женских особей.
5 в. Определение частоты встречаемости рецессивных фенотипов в мужской части популяции по сравнению с женской частью популяции Поскольку частоты двух гемизиготных генотипов в мужской части популяции при сцеплении с полом совпадают с частотами соответствующих аллелей у женских особей предыдущего поколения, то фенотипы, определяемые рецессивными генами, у мужских особей будут встречаться чаще, чем у женских. Если частота сцепленного с полом рецессивного аллеля равна , то частота определяемого им фенотипа будет равна для мужских особей и для женских особей.
 Отношение этих двух величин составляет: Чем меньше значение q, тем выше отношение частоты определяемого рецессивным геном фенотипа у мужских особей к его частоте у женских особей. Пример Частота рецессивного сцепленного с полом аллеля, вызывающего дальтонизм у людей (неспособность различать красный и зелёный цвета), составляет 0, 08; следовательно, этот дефект встречается у мужчин в 1/0, 08=12, 5 раза чаще, чем у женщин.
Отношение этих двух величин составляет: Чем меньше значение q, тем выше отношение частоты определяемого рецессивным геном фенотипа у мужских особей к его частоте у женских особей. Пример Частота рецессивного сцепленного с полом аллеля, вызывающего дальтонизм у людей (неспособность различать красный и зелёный цвета), составляет 0, 08; следовательно, этот дефект встречается у мужчин в 1/0, 08=12, 5 раза чаще, чем у женщин.