
Клеточные покровы и клеточная стенка грибов Клеточные покровы


Клеточные покровы и клеточная стенка грибов
 и полисахаридный чехол/капсула")
Клеточные покровы грибов= клеточная стенка (cell wall) и полисахаридный чехол/капсула
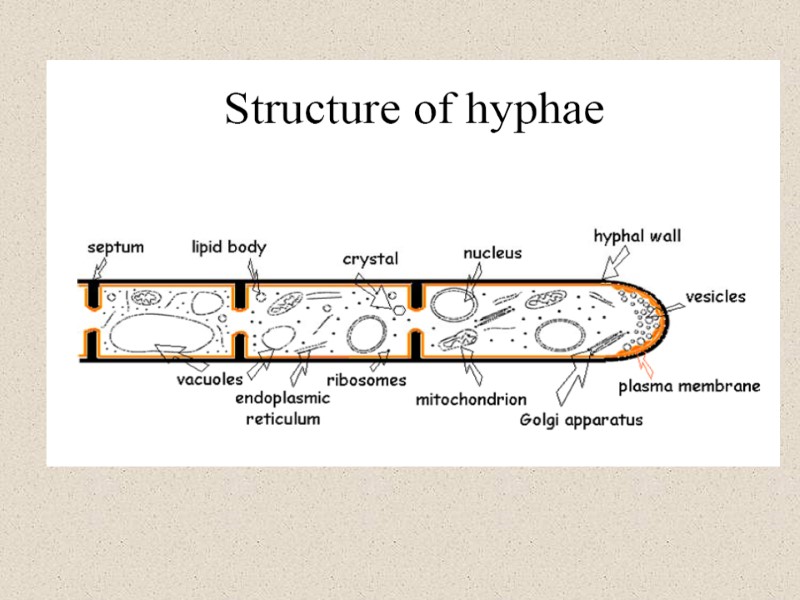
")
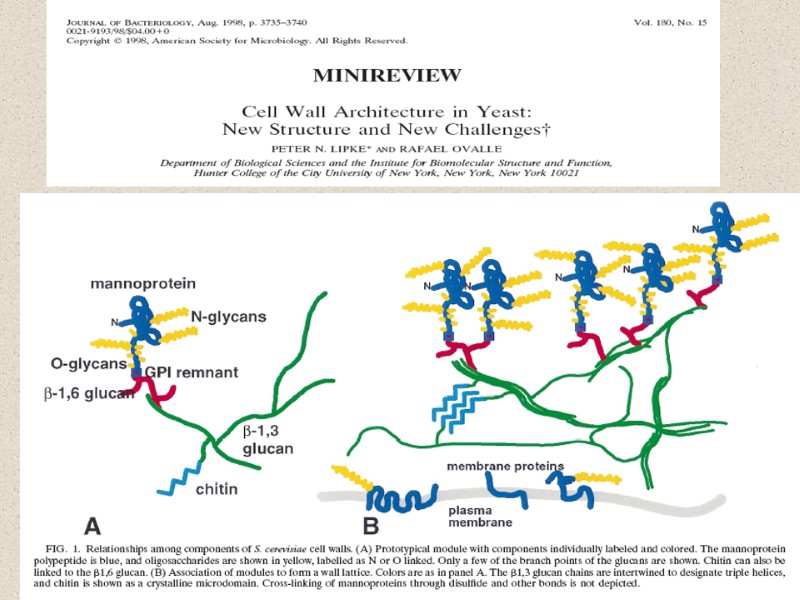
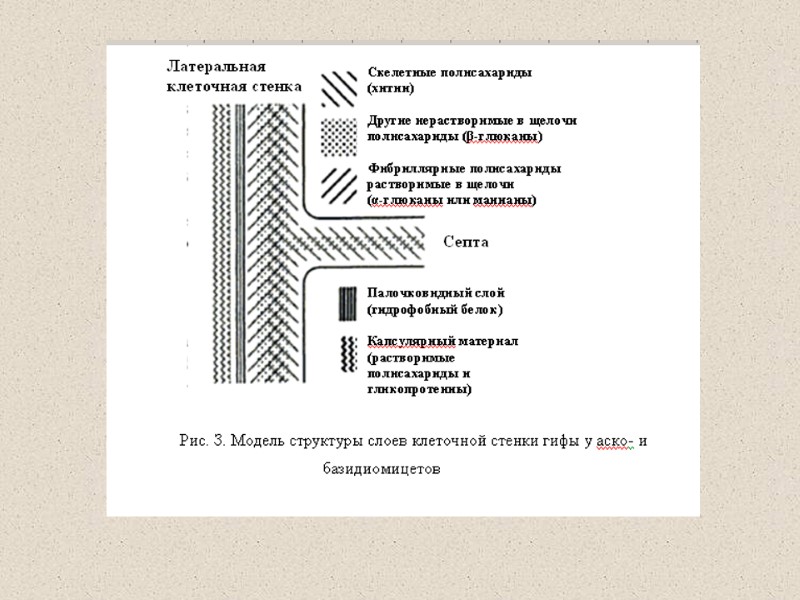
Рис. И. Схема строения клеточной стенки дрожжей GPI, glycophosphatidylinositol.(Selitrennikoff, 2001)
 Определяет морфологию клеток грибов. Сложно устроенные")
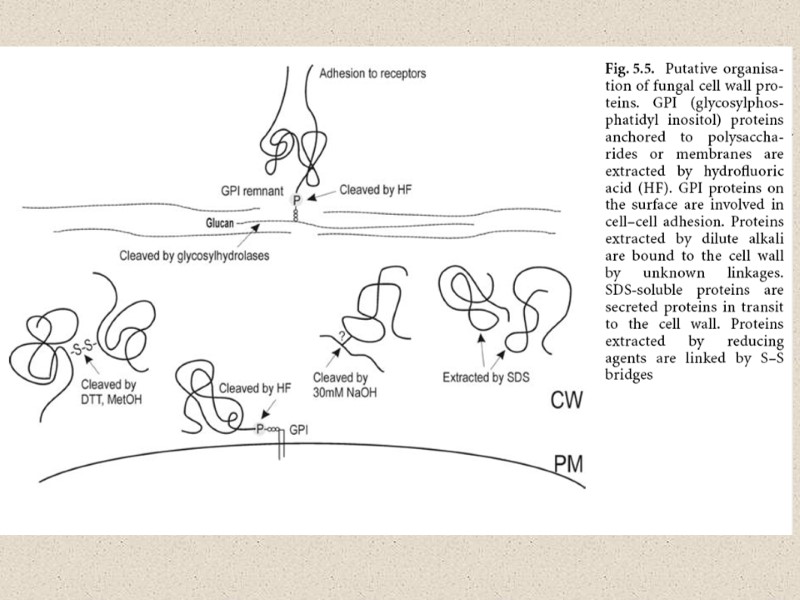
Функции клеточной стенки и клеточных покровов (КС) Определяет морфологию клеток грибов. Сложно устроенные многоклеточные плодовые тела высших грибов являются поразительным примером роли клеточной стенки в морфогенезе; Стабилизирует внутреннее осмотическое давление, что очень важно для сохранения формы клетки. Крепкая и эластичная стенка создает противоположное давление, которое останавливает поток воды извне; Защищает против физического стресса и функционирует как защитный покров. Комбинирование механической прочности и высокой эластичности позволяет стенке передавать и перераспределять физический стресс, обеспечивает эффективную защиту от разрушения/повреждения и позволяет гифе проникнуть и расти внутри агрессивного субстрата (Money, 2004; 2008); Клеточная стенка является скелетом/опорой для белков. Большая механическая прочность стенки обеспечивается полисахаридами, но они в тоже время выполняют функцию опоры для гликопротеинов наружного слоя, прикрепленных к ним. Функции гликопротеинов - ограничивать проницаемость стенки (в обоих направлениях), создавать микроокружение во внутренних слоях стенки смежных с мембраной, которая находится под контролем гриба. Отрицательно заряженные фосфатные группы на их углеводородной стороне цепи, вероятно, вносят вклад в удержание воды, а белки наружного слоя участвуют в узнавании: субстрата, партнеров при спаривании, субстратов – хозяев для патогенных грибов, последующей адгезии и захвата субстрата.

Методы изучения химического состава клеточных стенок
")
Методы изучения структуры световая микроскопия просвечивающая электронная микроскопия (иммуноцитохимия)
 Три главные группы: глюканы, маннаны и")
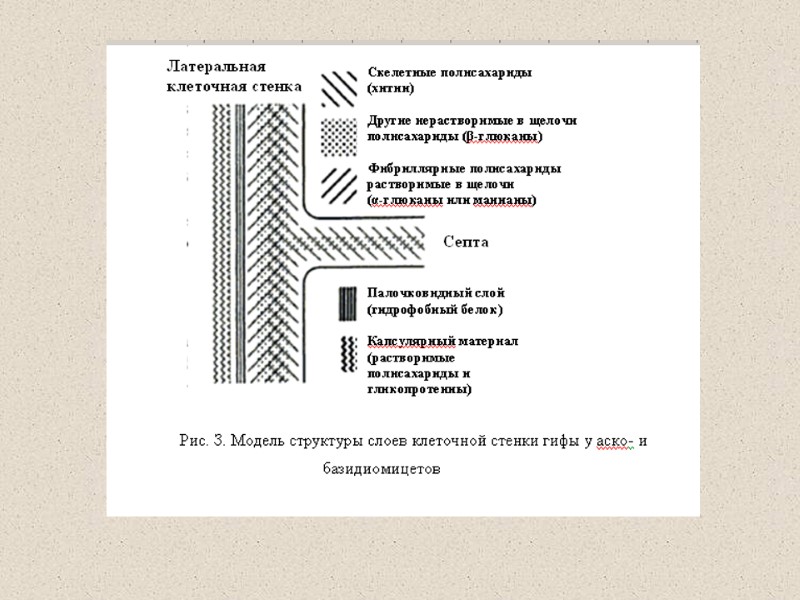
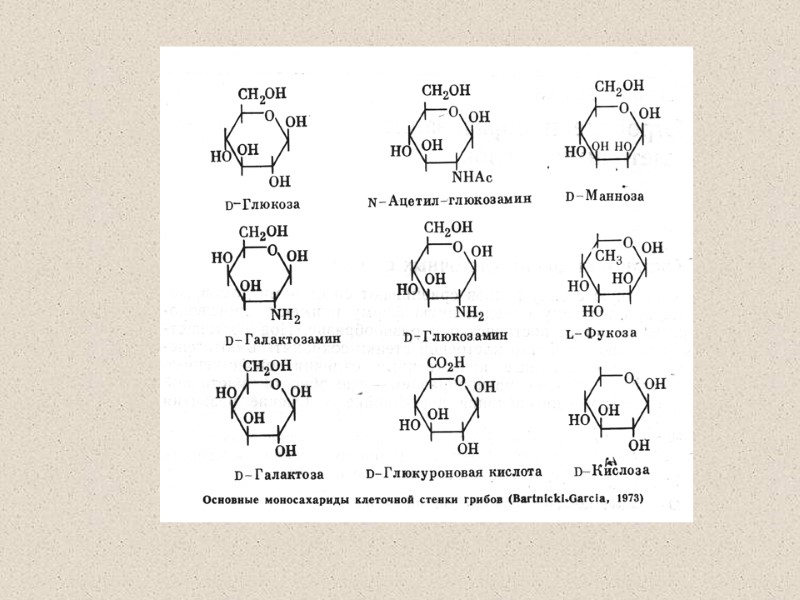
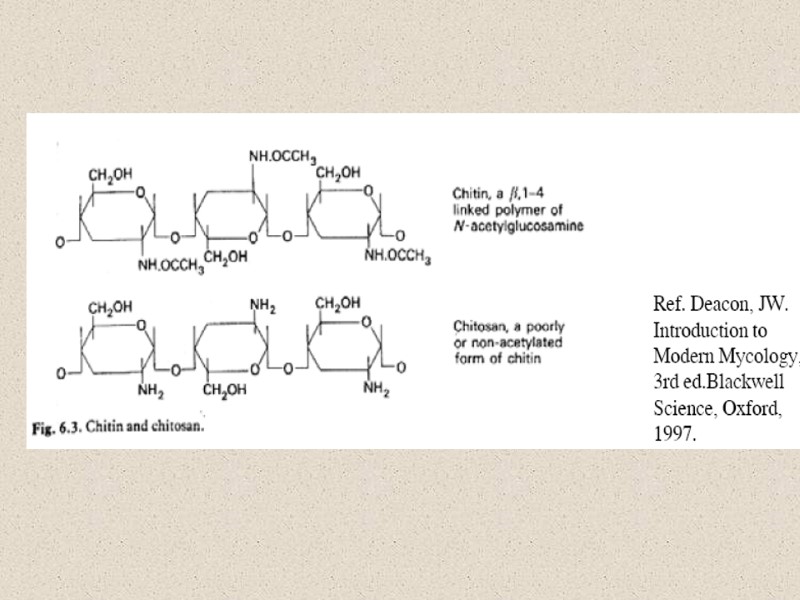
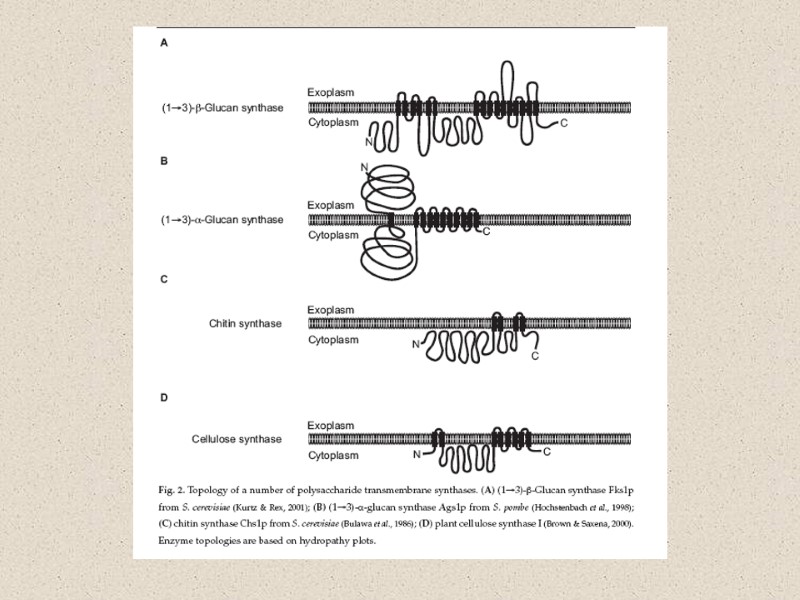
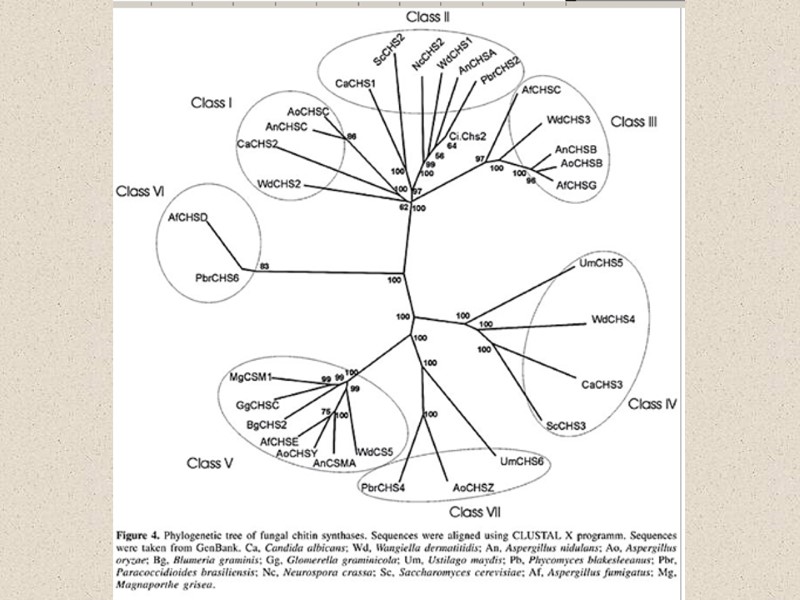
Компоненты КС (90% полисахариды) Три главные группы: глюканы, маннаны и хитин Два главных типа компонентов КС различаются по функциям: структурный компонент, который определяет жесткость КС компоненты матрикса – связующий материал между фибриллами различия по структуре структурные компоненты (щелоченерастворимы) хитин (полимеры β-1,4-связанных остатков N-ацетилглюкозамин); и хитозан (деацетилированный хитин) глюкан – полимеры β-1,3-связанных остатков глюкозы с короткими β-1,6-связанными боковыми веточками целлюлоза - β-1,4-связанные глюканы (очень редко) матриксные компоненты (растворимы в щелочи) глюкуроновые кислоты и др. маннопротеины (манноза связанная с белком), галактоманнопротеины и др.

целлюлоза
")
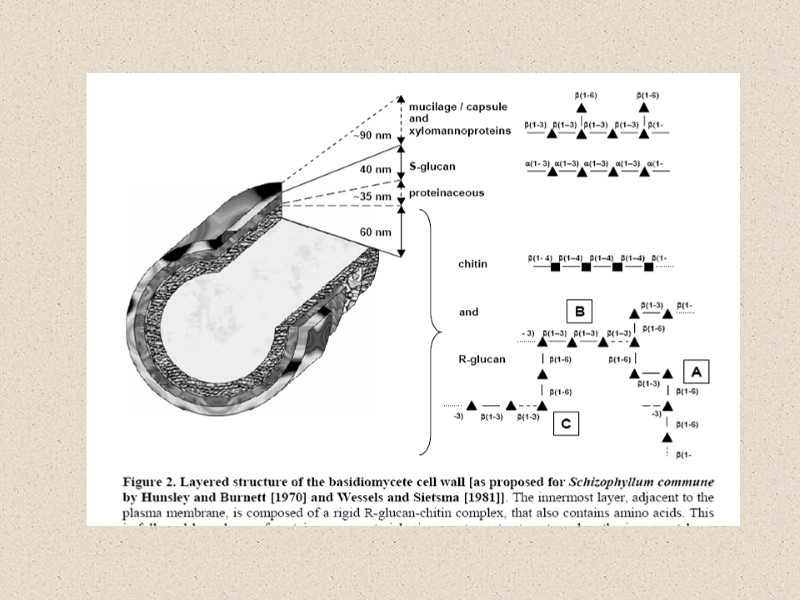
Молекулярная организация клеточной стенки у разных грибов (Howard and Gow, 2007)

Композиция клеточной стенки
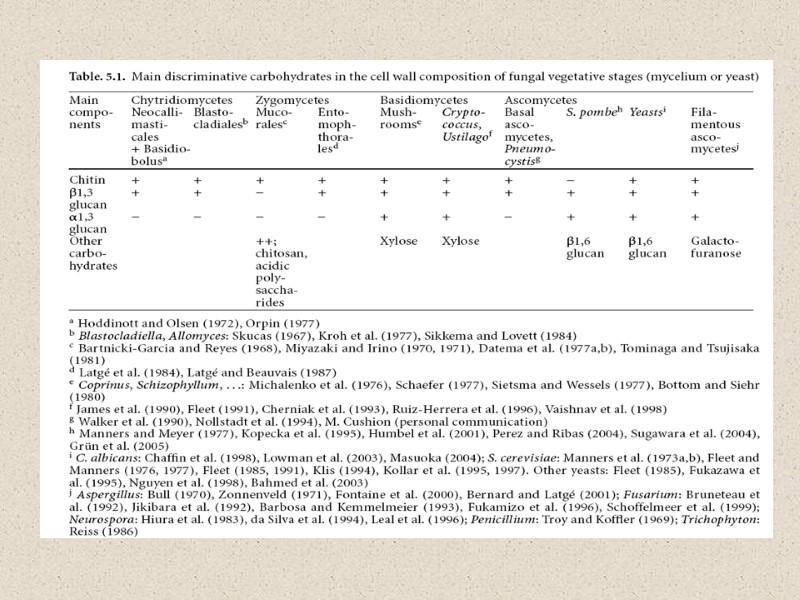
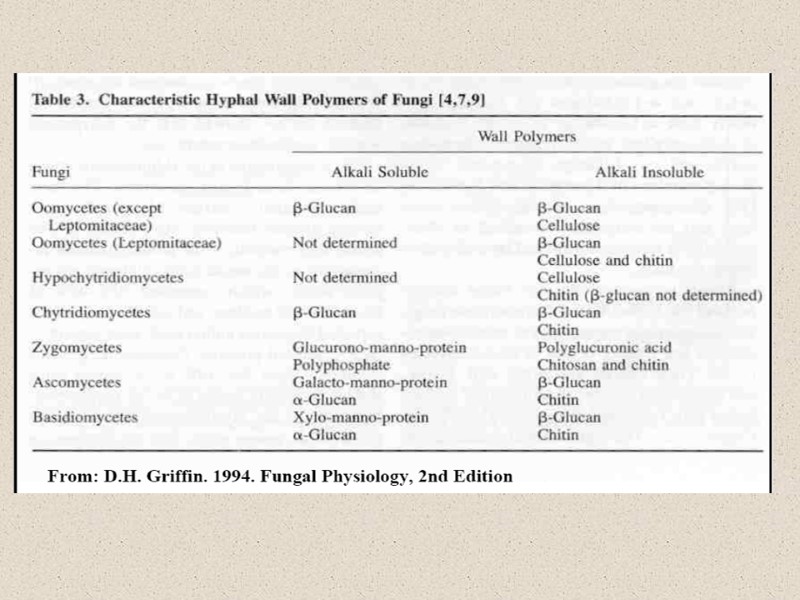

Эволюция клеточной стенки


Структурная и молекулярная организация клеточной стенки у разных грибов
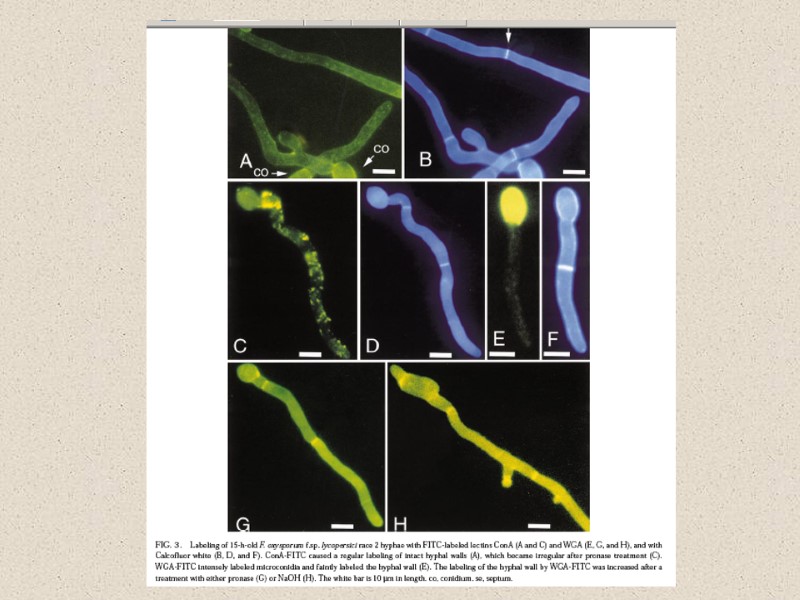
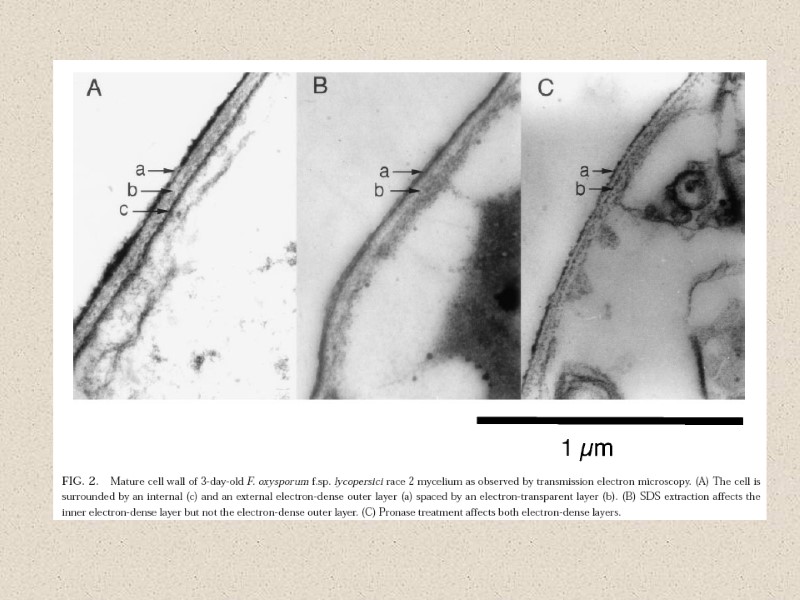

Факторы, влияющие на состав КС: -стадия жизненного цикла -условия культивирования

Electron microscopy of U. maydis cell sections. Median sections of cells stained with KMnO4. (A) Mycelial cell. (B) Yeast cell. Magnification bars, A, 0.05 μm; B, 0.15 μm. Notice the different structures of both cell walls. Mycelial wall (A) displays its layered appearance. Layer b appears more electron dense than layers a and c. The surface appears covered by a loose electron-dense layer (*). Yeast wall appears electron transparent, and only the surface loose layer is stained (*).
 и глубинный мицелий")
Влияние условий культивирования на композицию клеточной стенки Agaricus bisporus (поверхностный (А) и глубинный мицелий (Б) А Б

Отличия по структурным полисахаридам между дрожжевой и мицелиальной формой Blastomyces dermatitidis, % (Kanetsuna and Carbonell 1971)/Nemecek et al. (2006)


Композиция клеточных стенок у спор и вегетативных клеток Saccharomyces cerevisiae

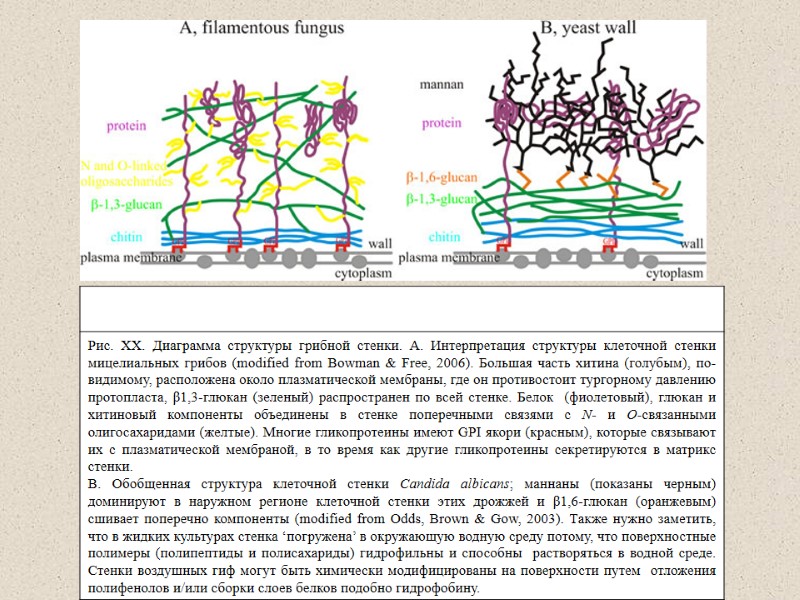
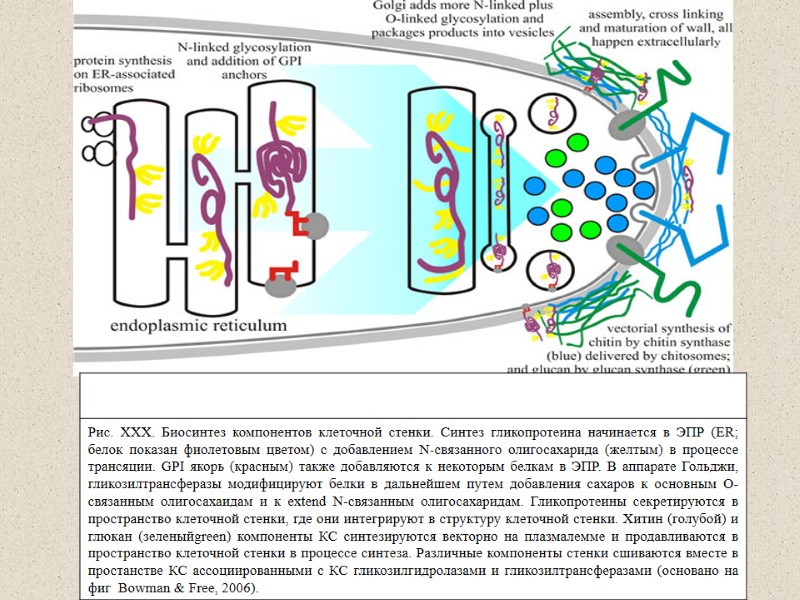
Representation of the fungal cell wall. The major components of the fungal cell wall arechitin, glucans and glycoproteins. Although species-specific variations exist, the cell wall components are thought to be arranged as shown. Most ofthe chitin is considered to be located near to the plasma membrane. The beta-1,3-glucan extends throughout the cell wall. The glycoproteins are extensively modified with N- and O-linked oligosaccharides. Many of the glycoproteins have GPI anchors (гликозилинозитолфосфатидил), which tether them to the plasma membrane while other glycoproteins are secreted into the cellwall space. The proteins, glucans and chitin components are integrated into the wall by crosslinking the chitin, glucans, protein-associated oligosaccharides and GPI anchors together. The structure and synthesis of the fungal cell wall Shaun M. Bowman and Stephen J. Free,2008

Механизм образования клеточной стенки
 during morphogenesis of yeasts and conidia")
Cell wall modifications (synthesis and lysis) during morphogenesis of yeasts and conidia

Мицелиально-дрожжевой диморфизм
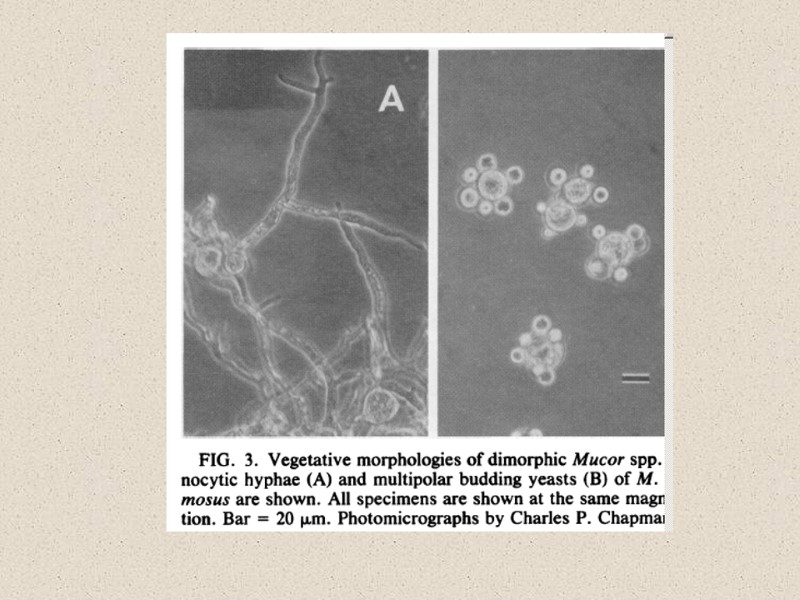
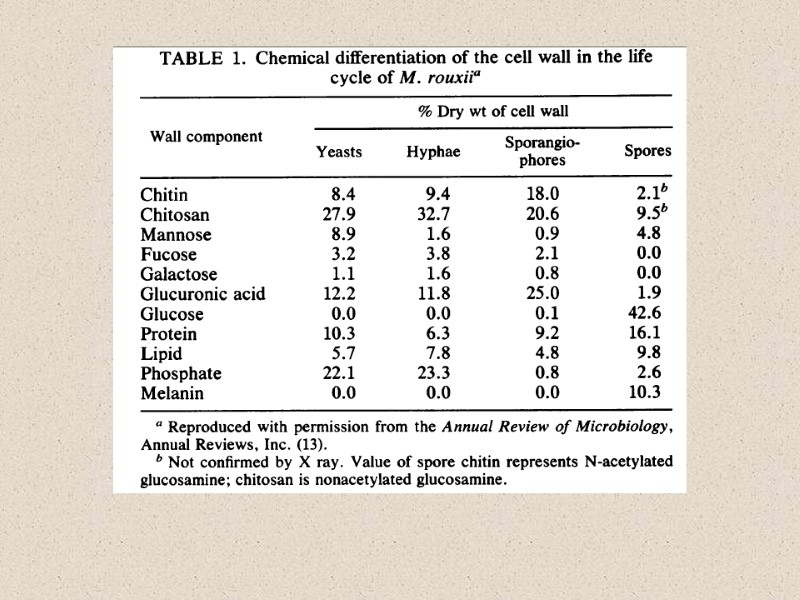
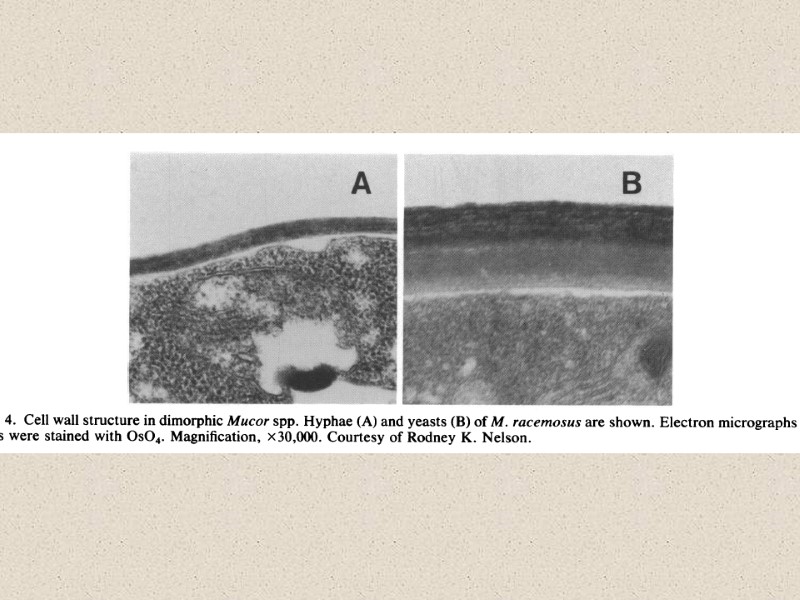
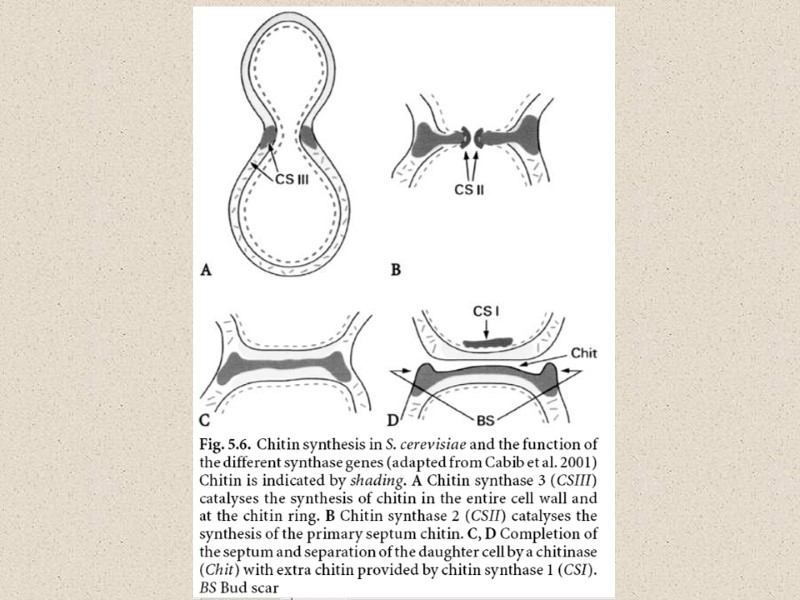
 Michalenko")
Очищенная фракция бесцветных клеточных оболочек вегетативного мицелия шампиньона содержит (% сух вес) Michalenko G.O., Hohle H.R., Rost D. Chemistry and architecture of the mycelial wall of Agaricus bisporus // J.Gen.Microbiol. 1976. V.92. № 2. P. 251-262. Таблица 3
/Nemecek et al. (2006) Химический состав клеточных стенок мицелия и")
(Kanetsuna and Carbonell 1971)/Nemecek et al. (2006) Химический состав клеточных стенок мицелия и дрожжей у Blastomyces dermatitidis

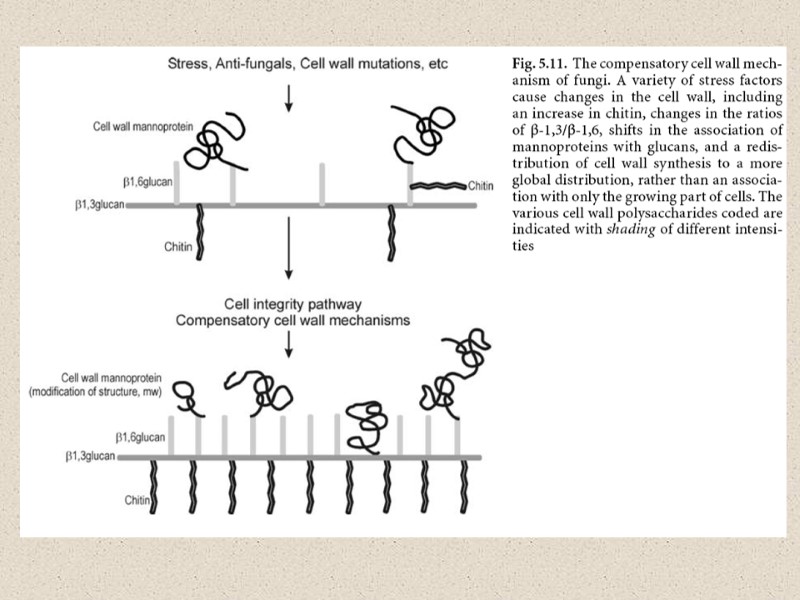
компенсаторные механизмы

1 2 3

Структурные белки, связанные с клеточной стенкой: гидрофобины помимо гидрофобинов важную роль в агрегации гиф и межклеточных контактах играют галектины (особый класс лектинов). Галектины животных вовлечены в процессы роста, дифференциации и апоптоза клеток (Сooper, 2002).

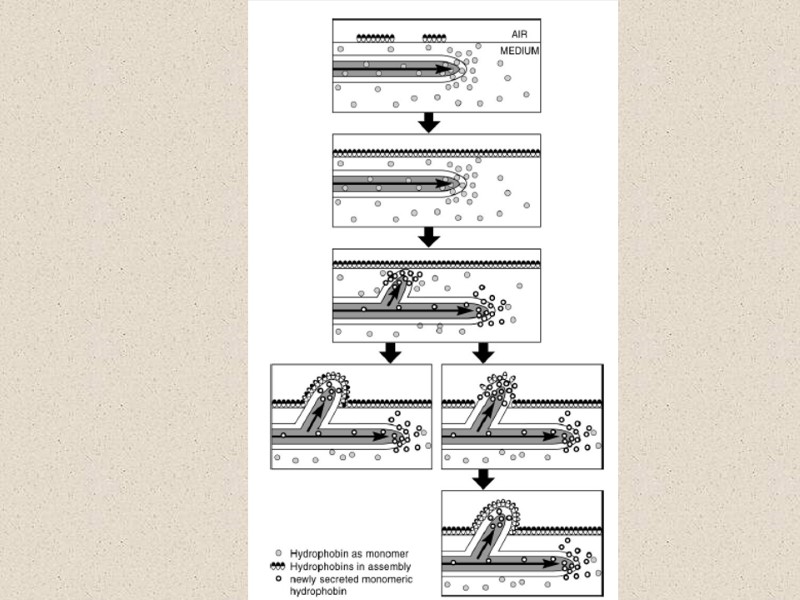
Гидрофобины – большая неоднородная группа родственных белков широко распространенных у мицелиальных грибов. Они образуются грибами в процессе их жизнедеятельности и могут составлять до 10% от общего белка стенки. Каждая молекула состоит из гидрофобного домена и гидрофильного домена. Амфипатичная структура обеспечивает экстраординарные функции в процессе роста и развития как элиситоры и токсины. Белки экскретируются на кончике гифы. Если гифа находится в водной среде, гидрофобины переходят в раствор. Если гифа выходит из раствора наружу, полипептид полимераза располагается параллельно поверхности гифы. Протеин оказывается повернутым к клеточной стенке гидрофильным концом, а гидрофобный конец направлен наружу. Такая структура снижает движение воды через клеточную стенку гифы, обеспечивая защиту от высыхания. А также увеличивает прочность стенки. Гидрофобный домен обеспечивает взаимодействие с гидрофобными поверхностями. Этими поверхностями могут быть другие гифы, что приводит к образованию структурного комплекса (плодовое тело). Гифы могут также взаимодействовать с гидрофобными поверхностями растений, способствовать взаимодействию спор перед формированием аппресориев. Гидрофобины выполняют важную роль в морфогенезе грибов. Показано, что гидрофобиновый домен у грибов – микобионтов лишайников контролирует движение воды по таллому. Домены обеспечивают защиту logging воды в воздушном пространстве, позволяет перемещение воды и питательных веществ по каналам, контролируемым грибом. Это позволяет предположить механизм регуляции извлечения (output) фотобионта that is essentially mechanistic. Каждый гриб имеет более одного гена и более 10 разных гидрофобинов. Разные гены гидрофобинов экспрессируются в разное время. У Schizophyllum белок, обнаруженный в стенках гифы отличается от белка, экспрессируется в трубочках (tubes) базидиокарпа. Гены экспрессируются по-разному у монокарионов и дикарионов одного и того же гриба. Регуляция экспрессии является частью комплекса регуляторных процессов, которые еще не достаточно изучены. Гидрофобины участвуют в широком спектре процессов. Особенно, где гифа контактирует с другими поверхностями, модификация движения растворов через стенку or require strength и ригидность мы можем обнаружить присутствие гидрофобинов.
 Tip of a conidium of Penicillium notatum, freeze-fractured. Bar, 500 nm. (B) Surface")
(A) Tip of a conidium of Penicillium notatum, freeze-fractured. Bar, 500 nm. (B) Surface of conidium of Neurospora crassa showing the EAS-encoded rodlet layer, freeze-fractured. Bar, 100 nm. (C) Surface of conidium of Aspergillus nidulans showing the rodA-encoded rodlet layer, freeze-fractured. (D) Surface of the pileus of Agaricus excellens mushroom showing rodlet layer. (E) Surface of conidium of M. grisea. Direct shadowing at room temperature to show fine structure of surface rodlets encoded by MPG1. Note the bootlace internal structure on rodlets. (F) Surface of a conidium of an mpg12 null mutant showing the absence of a rodlet layer. (Kershaw and Talbot, 1998)
/капсулы")
Чехлы (sheath)/капсулы
 M. Lars. et Lomb) – место локализации экстраклеточных")
Чехлы дереворазрушающих грибов (Postia placenta (Fr.) M. Lars. et Lomb) – место локализации экстраклеточных ферментов и грибных метаболитов Proposed functions for the extracellular sheath include (i) modification of the extracellular ionic environment and pH, (ii) recognition of and adhesion to the substrate, (iii) concentration, storage, and transport of decay agents, (iv) protection against dehydration and environmental injury, (v) conditioning of the woody substrate to degradation, (vi) storage of nutrients, especially glucose, (vii) regulation of the decay process, and (viii) increased surface area for aerobic respiration (Highley 1987; Eriksson 1988; Ruel et al. 1990)

Чехол у Laetiporus sulphureus

Чехол у Pleurotus pulmonarius выполняет важную роль в межклеточных контактах

Капсула дрожжей Соединения, образующие капсулу, могут относится к следующим группам: Фосфоманнаны – синтезируемые некоторыми аскомицетовыми дрожжами рода Pichia, образуют тонкий слой на поверхности клетки. α-Глюканы, к числу которых относится пуллулан, образуемый дрожжеподобным грибом Aureobasidium pullulans. β-Маннаны – образуются пигментированными дрожжами из родов Rhodotorula, Sporobolomyces. Гетерополисахариды – характерны для многих дрожжей из родов Cryptococcus и Lipomyces. Функции капсулы дрожжей: -прикрепление клеток к поверхности твердого субстрата, -резервный источник питания, -улучшение водного снабжения клетки (в том числе защита от высыхания) -в природных местах обитания – места аккумуляции спутников и ассоциантов грибов.
, forming up to 88% of")
Most capsular mannose exists as the polysaccharide glucoronoxylomannan (GXM), forming up to 88% of capsular material (Cherniak and Sundstrom 1994). Капсула Cryptococcus neoformans играет главную роль в вирулентности

Кристаллы оксалата кальция на поверхности клеток мицелия Agaricus bisporus выделение излишних ионов Са2+ и солей щавелевой кислоты
 TEM of C. neofomans TEM of (a) melanin-lacking")
Меланин (защита от противогрибных препаратов) TEM of C. neofomans TEM of (a) melanin-lacking (500w) and (b) melaninproducing (500b) C. neofomans cells grown in L-DOPA medium for 7days. Similar results were obtained with the other two pars of isolates. Scale bar, 500nm, (Magnification X 15000). a b

Модель структуры меланина у C. neofomans
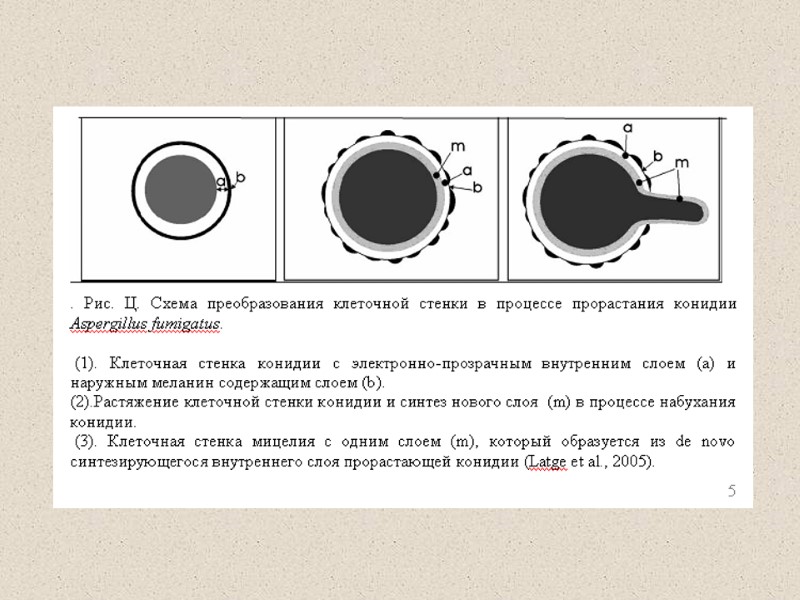
 of the conidium (c–e) of A. fumigatus.")
Ultrastructural features of the cell wall (cw) of the conidium (c–e) of A. fumigatus. a, c, Scanning electron micrographs; d, transmission electron micrographs of sections of the conidial cell wall (note the dense melanin outer layer in the conidial wall); e, back-scattered electron micrograph of a conidium labelled with laminin, anti-laminin antibody and protein A-gold particles (arrow heads) (courtesy of G. Tronchin); f, carbon–platinum replica of the conidial surface showing the rodlet structure.

Функции меланина: защита от токсинов устойчивость к экстремальным температурам устойчивость к ультрафиолетовой радиации защита от нематод и амеб повышает резистентность к факторам иммунной системы снижает восприимчивость к антигрибным препаратам
20248-cell_wall_lekciya_1-2012.ppt
- Количество слайдов: 64