
Клеточная стенка грибов Грибная стенка — рабочая органелла


Клеточная стенка грибов

Грибная стенка - рабочая органелла Грибная клеточная стенка является важным филогенетическим и таксономическим признаком, но она является динамичной органеллой, которая имеет четыре главные функции: Определяет форму клетки. Стенка определяет морфологию гифы или другой клетки гифы. Наиболее эффектное многоклеточные плодовые тела высших грибов поразительный пример как клеточная стенка вносит вклад в морфогенез; Стабилизирует внутреннее осмотическое давление это очень важно для сохранения формы клетки. Стенка крепкая и эластичная и создает противоположное давление, которое останавливает поток воды; Защищает от физического стресса, и функционирует как защитный покров. Комбинирование механической прочности и высокой эластичности позволяет стенке передавать и перераспределять физический стресс, предлагают эффективную защиту против механической опасности и позволяют гифе проникнуть и расти внутри агрессивного субстрата (Money, 2004; 2008); Клеточная стенка как опора для белков. Большая механическая прочность стенки обеспечивается ее переносящими стресс полисахаридами, но они также выполняют функцию опоры для гликопротеинов наружного слоя, прикрепленных к ним. Функции этих гликопротеинов - ограничивать проницаемость стенки (в обоих направлениях), создавать микроокружение во внутренних слоях стенки смежных с мембраной, которая находится под контролем гриба. Отрицательно заряженные фосфатные группы на их углеводородной стороне цепи вероятно вносят вклад в удержание воды, а белки наружного слоя участвуют в узнавании партнеров спаривания, субстрата, субстрата и хозяев, и последующей адгезии и захвата какого-нибудь или всех из них.

Структура клеточной стенки Клеточная стенка имеет структурный компонент (собственно клеточная стенка, расположена снаружи от плазмалеммы) и внеклеточный матрикс. Структурно, грибная стенка трехмерная сеть из полисахаридов, гликопротеинов и белков, многие из которых уникальны для грибов. Типичные стенки содержат фибриллярные полисахариды высоко эластичные прочные погруженные в более геле-подобный матрикс, содержащий разнообразные полисахариды, гликопротеины и белки с разнообразными минорными компонентами, включающими липиды, пигменты, неорганические ионы и соли. Фибриллярный материал стенки высоко инертен, образуя главный структурный компонент, но композиция других материалов изменяется со временем и положением так как эти компоненты могут служить как: питательные резервы, в транспорте и передвижении, для метаболизма непроницаемых веществ, для коммуникаций и взаимодействия с внешней средой, для защиты от наружных атак.

Внеклеточный матрикс –компонент клеточной стенки грибов


Белки Протеины редко составляют более 20% материала стенки, и большая часть это гликопротеины. Некоторые белки выполняют структурную роль, но большинство вносят вклад во многие другие функции упомянутые выше. Белки у или около наружной поверхности определяют поверхностные свойства стенки: то есть, или она гидрофобная или гидрофильная (т.е несмачиваемая или смачиваемая) или связывающая или антигенная. Низкие концентрации липидов и восков в грибных стенках обычно служат для контроля движения воды, особенно при защите от высыхания.

Полисахариды Примерно 80% компонентов стенки составляют полисахариды. В гифе главный компонент стенки и наиболее важный структурный компонент это хитин, который является полимером β1,4-связанных N-ацетилглюкозамина

Ковалентно связанные структура N-ацетилглюкозамина и его гомополимера хитина, который синтезируется ферментом хитинсинтазой. Природный хитин имеет молекулярную массу несколько млн. Процесс экстракции фрагментирует полимер и молекулярная масса коммерческого препарата хитина варьирует от 350000 до 650000

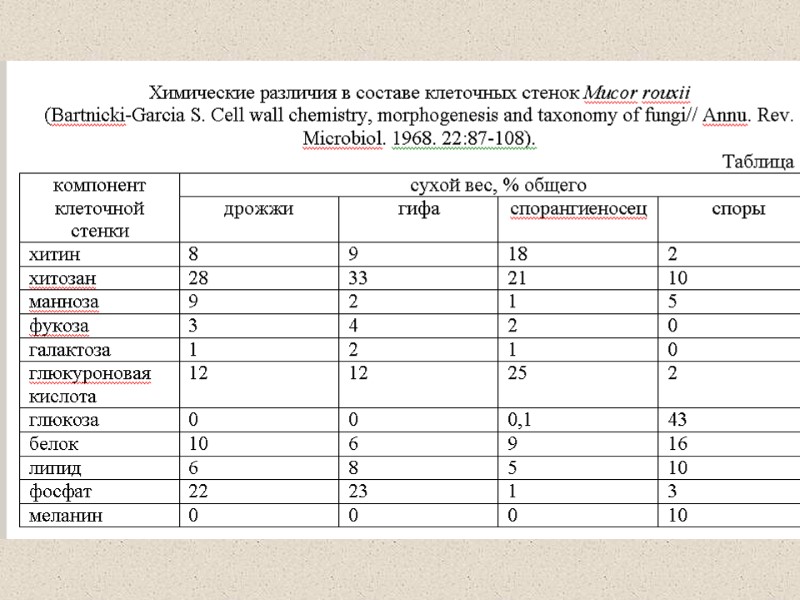
Содержание хитина у разных грибов 1-2% хитина от сухого веса КС Aspergillus Neurospora дрожжи 10 – 20 % от сухого веса КС

Полисахариды клеточной стенки

Различия в химическом составе полисахаридов у разных таксонов

Различия в химическом составе полисахаридов у разных таксонов -клеточные стенки традиционно известных Zygomycota содержат хитин и хитозан (полимер глюкозамина образующегося из хитина путем удаления N-ацетильных групп) среди более волокнистых полимеров и полиглюкуроновая кислота и глюкурономаннобелки среди водно-растворимых гельподобных полимеров. -Ascomycota и Basidiomycota оба содержат хитин вместе с β1,3, β1,6-связанных глюканов в фибриллярной форме, но Ascomycota содержат гелеподобный α1,3 глюкан и галактоманнопротеины, у Basidiomycota ксиломаннопротеины обнаружены поблизости от α1,3 глюкана (Gooday, 1995a, Bowman and Free, 2006).


Появление полисахаридов в КС грибов, основанное на таксономических кластерах к временной геологической шкале. Длина ветвей пропорциональна средней последовательности ядерной SSU rDNA (стрелка ↓ появление полисахарида, стрелка ↑утрата) (Latge, 2007).

заключение - главными структурными веществами стенки являются полисахариды, в большинстве своем – глюкан, и у мицелиальных грибов определяющий форму компонент – хитин; - различные полисахаридные компоненты связаны вместе водородными связями и ковалентными связями; - множество белков и гликопротеиновых компонентов вносят вклад в функцию стенки; некоторые из них структурные, часть - ферменты, часть имеют разнообразные биологические и биофизические характеристики на наружной поверхности клеточной стенки; - белки могут быть заякорены в плазматической мембране, ковалентно связываться с полисахаридами стенки или более тесно ассоциировать со стенкой; - стенка – это динамичная структур,а которая модифицируется: с возрастом и/или как часть дифференциации гифы и/или при кратковременных изменениях физических или физиологических условий

Архитектура клеточной стенки
 и фрагмент мицелия (б)")
Архитектура клеточной стенки зависит от стадии жизненного цикла, Спора (а) и фрагмент мицелия (б) Agaricus bisporus U3 Многослойные клеточные покровы. У клеточных стенок мицелия исследованных аскомицетных и базидиомицетных грибов клеточные стенки в большинстве своем - двухслойные, реже однослойные и многослойные.

от условий культивирования
 Клетка мицелия.")
Электронная микроскопия срезов клеток U. maydis. Медианные срезы клеток окрашены KMnO4. (A) Клетка мицелия. (B) Дрожжевая клетка. Масштабные отрезки A, 0.05 μm; B, 0.15 μm. Мицелиальные стенки (A) демонстрируют слоистость. Слой b более электронно-плотный, чем слои а и с. Поверхность покрыта неплотно прикрепленным электронно-плотным слоем (*). от ядерного статуса…
 . Очень")
Строение КС Candida. albicans на срезах с низкотемпературной фиксацией (ТЕМ) . Очень деликатный фибриллярный внеклеточный слой (в виде щетки) покрывает наружную поверхность клеток. Фибриллярные слои ограничены границей раздела со средними слоями. Электрон-плотный материал остается между фибриллами (стрелка). В цитоплазме четко видны рибосомы, округлые или овальные ядра и митохондрии x 40,000
 картина поперечного среза клетки")
Модель клеточных покровов и структуры меланина Cryptococcus. neoformans. (A) картина поперечного среза клетки C. neoformans. Полисахаридная капсула (Cap.) окружающая клетку. Меланин (черным) локализован в клеточной стенке (CW), за плазмалеммой (PM). (B) Увеличенное строение меланинового слоя в клеточной стенке C. neoformans. Меланин составляет гранулы расположенные в несколько слоев. Согласно модели маленькие молекулы питательных веществ (сахара и аминокислоты) могут проходить через этот слой. Антитела связываются с меланином блокируя транспорт питательных веществ. Крупные молекулы лекарственных препаратов также не проходят в клетку.

Модель структуры клеточной стенки гифы у аско- и базидиомицетов

Схема строения клеточной стенки дрожжей

Диаграмма структуры грибной стенки
 клеточной стенки грибов Связанные с глюканом : - белки, содержащие GPI (гликозилфосфатидил")
Белки (20%) клеточной стенки грибов Связанные с глюканом : - белки, содержащие GPI (гликозилфосфатидил инозитол) якорь (гликопротеины-маннопротеины, галактоманнопротеины), на поверхности они вовлечены в межклеточную адгезию, спаривание. - ‘белки с внутренними повторами’, или (PIR)-белки, сшивающие β1,3-глюкановые цепи через эфирную связь, формируют белково-полисахаридный комплекс β1,3-глюкан–PIR- β1,3-глюкан и найдены повсюду во внутреннем слое стенки (Yin et al., 2007). – секреторные белки - цитоплазматические, митохондриальные и др. белки

Хитин, синтез Синтез хитин осуществляется хитин синтазой, интегрированным в мембрану ферментом, который катализирует перенос N-ацетилглюкозамина с уридин дифосфат (UDP)-N-ацетилглюкозамина к растущей цепи хитина

Большинство грибов имеет несколько генов хитин синтазы: от одного у Schizosaccharomyces pombe до десяти у некоторых мицелиальных грибов таких как Aspergillus fumigatus. Хитин синтаза является интегральным мембран-связанным ферментом (трансмембранный белок), транспортируется в микровезикулах (хитосомы) к местам активного синтеза клеточной стенки.

Разные изозимы хитин синтазы могут быть связаны с разными специфичными функциями, которые зависят от времени и точки развития такие, как формирование септы и спор у Aspergillus (Ichinomiya et al., 2005), аппрессория у Colletotrichum (Werner et al., 2007), для ремонта или ответа на стресс (Bowman & Free, 2006). Некоторые могут даже быть важными для патогенности некоторых видов, мутанты с делецией не патогенны –признак, который показывает критическое значение целостности клеточной стенки в инфекционном процессе у грибов (Werner et al., 2007; Martín-Urdíoz et al., 2008). Но наиболее важная функция которую вносит хитин синтаза – вклад в растяжение гифы
 Локализация двух хитин синтаз у нейроспоры")
Riquelme et al. (2007) Локализация двух хитин синтаз у нейроспоры
.")
Хитин синтазы дрожжей Эти дрожжи имеют три хитин синтазы (Chs1, Chs2, and Chs3). Chs3 требуется для формирования хитиновых колец (рубец) в основании развивающихся почек и для синтеза хитина в латеральной клеточной стенке в процессе вегетативного роста. Отвечает за синтез 80-90% всего хитина. Chs2 синтезирует хитин в первичной септе. Chs1 хитин синтаза реставрирует стенку, пополняя утраченные полимеры хитина в процессе цитокинеза.

Глюкан, синтез Хотя хитин обеспечивает механическую прочность стенки, глюкан составляет до 50–60% от сухого веса у мицелиальных грибов и от 65% до 90% клеточной стенки глюкан это β-1,3-глюкан. У грибов в клеточных стенках обнаружены глюканы с разными типами связи включая β-1,6, смесь β-1,3 с β-1,4, α-1,3, и α-1,4-связанными глюканами. Однако, β-1,3-glucan служит главным структурным элементом, с которым ковалентно связаны другие компоненты КС. Для формирования КС и нормального развития грибов требуется синтез β-1,3-глюкана (Latgé et al., 2005; Bowman & Free, 2006; Lesage & Bussey, 2006).

Как и в случае с хитином, полимеры глюкана формируются мультиединицами ферментного комплекса в плазматической мембране, полимер выбрасывается во внеклеточное пространство через векторный синтез. Линейный полимер синтезируется ферментом комплекса известным как β(1,3)-глюкан синтаза (Douglas, 2001; Latgé et al., 2005). Полисахаридные полимеры образуют цепочки длиной до 1500 глюкозных остатков; он становится нерастворимым с увеличением степени полимеризации, фермент прикрепляется к рождающемуся глюкану. Синтез векторный, новые глюкановые цепи сразу же выбрасываются во внеклеточное (периплазмтическое) пространство рядом с цитоплазматической мембраной. Это способствует их встраиванию в клеточную стенку в местах активного синтеза КС. Глюкан синтазные комплексы подобно хитин синтазам главным образом локализованы в районах активного растяжения, почкования, ветвления или септирования.

Гены кодирующие белок β-1,3-глюкан синтазу были впервые идентифицированы у Saccharomyces cerevisiae, которая содержит две каталитические субъединицы и один регуляторный белок Rho1 . Каталитические белки S. cerevisiae кодируются генами FKS1 и FKS2, каждый из них важны для формирования нормальной стенки. Разрушение или FKS1 или FKS2 гена приводит у мутантов к медленной скорости роста и дефектам клеточной стенки; одновременная делеция двух генов летальна.

Гены FKS и RHO1 высоко консервативны у других грибов. Геномы A. fumigatus и N. crassa содержат один ген каталитической субъединицы и один кодирует GTPase регуляторную единицу; оба гена необходимы для жизнеспособности клетки (Latgé et al., 2005).

Гликопротеиновый компонент В клеточной стенке грибов ветвящиеся глюканы связаны вместе поперечными связями с хитином, в дополнении глюкан имеет маннопротеины (дрожжи) (De Groot, Ram & Klis, 2005; Klis, Boorsma & De Groot, 2006). В клеточной стенке дрожжей (S. cerevisiae and C. albicans) белки достигают 30-50% от сухого веса стенки; В КС гиф N. crassa примерно 15% белка от сухого веса. У большинства изученных мицелиальных грибов КС содержат примерно 20-30% белка.

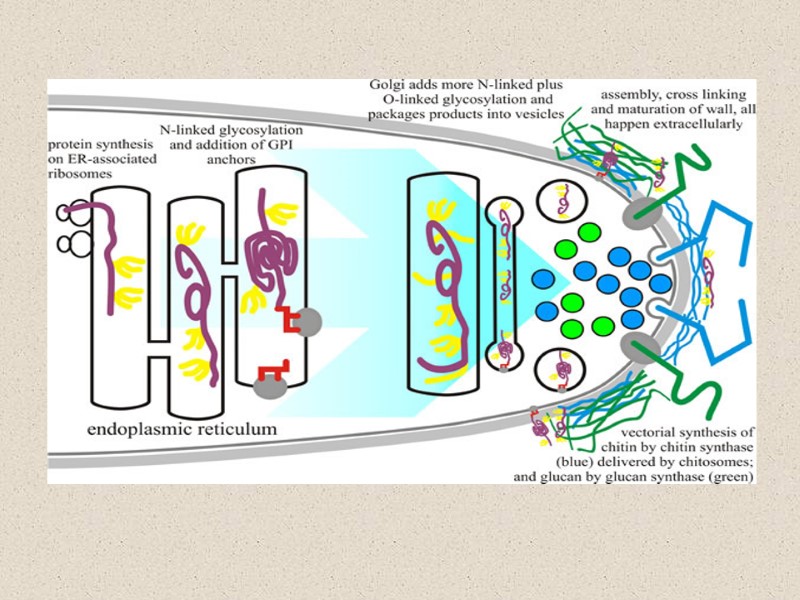
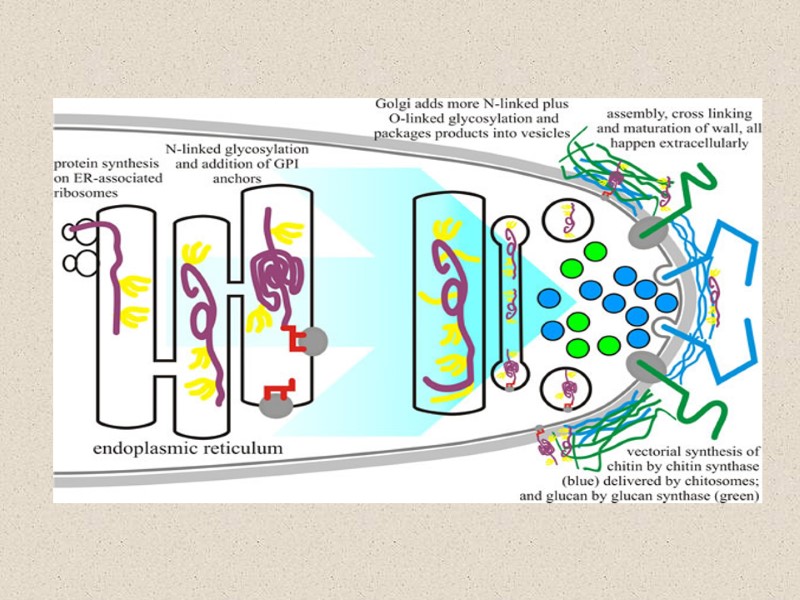
Синтез гликопротеина

Большинство белков клеточной стенки структурно интегрированы в стенку, будучи связанными с хитином и/или глюканом через сахара в N- и O-связанными олигосахаридами и/или с GPI якорем. ‘Белки с внутренними повторами’, или (PIR)-белки, сшивающие β1,3-глюкановые цепи через эфирную связь, формируя белково-полисахаридный комплекс β1,3-глюкан–PIR- β1,3-глюкан и найдены повсюду во внутреннем слое стенки (Yin et al., 2007).

Белки в клеточной стенке вносят вклад в форму клетки, адгезию, защиту против окружающего стресса, абсорбцию молекул, передачу внутриклеточных сигналов и восприятие внешних стимулов, а также синтез и ремоделирование компонентов стенки (Adams, 2004; De Groot et al., 2005; Klis et al., 2006). Интересная группа стеночных белков. Это цитоплазматические (даже митохондриальные) белки, такие как гликолитические ферменты. Раньше считали их контаминацией препаратов клеточных стенок грибов, но сейчас накоплены доказательства в пользу того, что они реально являются резидентными белками, поперечно-связанные с клеточной стенкой. Имеется предположение, что такие белки могут быть «органеллами стенки» с широким районом ферментных метаболических функций.

заключение Компоненты КС глюкан и хитин синтезируются векторно на цитоплазматической мембране и выдавливаются в пространство снаружи от плазмалеммы в процессе их синтеза. Гликопротеины стенки проходят через секреторные пути эндоплазматический ретикулум-Гольджи и секретируются в то же самое пространство стенки, где они интегрируются в структуру КС. Разные компоненты КС связываются поперечно на месте при участии ассоциированными с КС гликозилгидролазами и гликозилтрансферазами

Синтез стенки и ремоделирование Обобщенная молекулярная модель роста гифы

Процессы синтеза и ремоделирования происходят при: дифференциации гиф и спор, ветвлении гиф, септировании, формировании анастомозов

Ремоделирование зависит от координированной активности нескольких гликопротеинов уже присутствующих в структуре стенки. ‘Ассоциированные со стенкой ферменты’ вовлекают хитиназы, глюканазы и пептидазы (Cohen-Kupiec & Chet, 1998; Adams, 2004; Seidl, 2008); ферменты, которые осуществляют гидролиз и разрушение компонентов клеточной стенки, а также глюкозилтрансферазы, которые вовлечены в синтез и сшивку стеночных полимеров. Ферментная активность этих белков должна быть четко контролируема, так чтобы зрелая стенка имела достаточную эластичность (растяжение), и жесткость, предохраняя от клеточного лизиса. Несколько ассоциированных со стенкой ферментов было идентифицировано у S. cerevisiae и A. fumigatus и показано участие в ремоделировании стенки (Bowman & Free, 2006).

Вторичное утолщение КС мицелия плодовых тел Schizophyllum commune за счет отложения глюкана щелоче-нерастворимый компонент клеточной стенки, который был назван R-глюкан

Вторичное утолщение КС мицелия шампиньона двуспорового

Пигменты КС Меланины - низкомолекулярные фенольные соединения. Каротиноиды -тетратерпены и тетратерпеноиды
 картина поперечного среза клетки")
Модель клеточных покровов и структуры меланина Cryptococcus. neoformans. (A) картина поперечного среза клетки C. neoformans. Полисахаридная капсула (Cap.) окружающая клетку. Меланин (черным) локализован в клеточной стенке (CW), за плазмалеммой (PM). (B) Увеличенное строение меланинового слоя в клеточной стенке C. neoformans. Меланин составляет гранулы расположенные в несколько слоев. Согласно модели маленькие молекулы питательных веществ (сахара и аминокислоты) могут проходить через этот слой. Антитела связываются с меланином блокируя транспорт питательных веществ. Крупные молекулы лекарственных препаратов также не проходят в клетку.

Белковые компоненты наружной стороны КС Агглютинины Адгезины Гидрофобины
 между клетками разных типов спаривания у Saccharomyces")
Агглютинины Агглютинины ответственны за ‘агглютинацию’ (межклеточную адгезию) между клетками разных типов спаривания у Saccharomyces cerevisiae на ранних этапах в спаривании. Взаимодействие имеет место между двумя заякоренными с наружной стороны плазматической мембраны двух спаривающихся клеток GPI-anchored гликопротеинами.
, а также в многоклеточной")
Адгезины Адгезины осуществляют гомотипичные взаимоотношения в спаривании (у почкующихся дрожжей), а также в многоклеточной дифференциации, например, известны гликопротеины (галектины), которые специфичны для плодового тела Coprinopsis. Адгезины осуществляют также адгезию к поверхностям. Фибриллы распространяются от стенки в суспензию жидкости обычно находят там, где грибы прикасаются к поверхностям. Фибриллы высоко специфичны к комплементарной молекулярной структуре на поверхности. Эта система узнавания общения между грибом и партнером широко распространена у патогенных и мутуалистических взаимоотношений, содействуя прикреплению к поверхности организмов хозяев (животное и растение). Но они также содействуют формированию биопленок, как на природных поверхностях и на катетерах и других медицинских приборах.
 Обнаружены у аско-")
Гидрофобины уникальные белки грибов (до 10% от общего белка КС) Обнаружены у аско- и базидиомицетов (7-9 kDa, двухдоменные белки ) амфифильные Модель формирования грибных воздушных структур после погружения

Функции гидрофобинов Способность гифы преодолевать поверхность раздела воздух/вода во влажных местах обитания; Обеспечивает гидрофобностью гифы и споры при контакте с воздухом; Участвуют в морфогенетических сигналах, инициирующих конидиеобразование и формирование плодовых тел; Участвуют в формировании ткани, особенно в контролировании обмена жидкость-воздух; Обеспечивают адгезию между гидрофильными клеточными стенками гриба и гидрофобными поверхностями растений и насекомых и поэтому способствует инфицированию и внедрению в поверхность хозяина; *интересно, воздушные конидии Aspergillus, Penicillium и Cladosporium которые имеют поверхностный слой гидрофобина не активируют иммунную систему (Aimanianda et al., 2009); Опосредует симбиотические взаимоотношения с корнями растений (микориза) и водорослями (лишайники).

Гидрофобины выделяются из кончика гифы; если гифа находится в водном окружении, гидрофобины проходят в раствор. Но молекулы белка способны к самосборке в покровной пленке границы вода/воздух (= гидрофильные/гидрофобные) и когда гифа появляется из раствора, полипептид полимеразы на поверхности стенки гифы, формирует пучки параллельных палочек. Каждая молекула гидрофобина связана с грибной стенкой гидрофильным концом; гидрофобный домен обращен наружу (Linder et al., 2005; Cox & Hooley, 2009). По растворимости гидрофобины делят на две группы: класс I гидрофобины формируют трудно растворимые мембраны, которые расщепляются только трфторуксусной кислотой и formic муравьиной кислотой, в то время как гидрофобины класса II растворимы охотно в этаноле или SDS.
 палочковидного слоя (A) Tip of a conidium of Penicillium notatum,")
Ультраструктура гидрофобного (класс 1) палочковидного слоя (A) Tip of a conidium of Penicillium notatum, freeze-fractured. Bar, 500 nm. (B) Surface of conidium of Neurospora crassa showing the EAS-encoded rodlet layer, freeze-fractured. Bar, 100 nm. (C) Surface of conidium of Aspergillus nidulans showing the rodA-encoded rodlet layer, freeze-fractured. (D) Surface of the pileus of Agaricus excellens mushroom showing rodlet layer. (E) Surface of conidium of M. grisea. Direct shadowing at room temperature to show fine structure of surface rodlets encoded by MPG1. (F)Surface of a conidium of an mpg12 null mutant showing the absence of a rodlet layer. (Kershaw and Talbot, 1998)

Гликопротеин гломалин Образуется в клеточных стенках гиф и спор арбускулярно микоризных грибов и накапливается в почве и около корней.

Минорными компонентами клеточных покровов являются липиды, меланин и оксалат кальция (на поверхности клеток мицелия Agaricus bisporus) Нейтрализация щавелевой кислоты, накопление ионов кальция
/капсулы")
Чехлы (sheath)/капсулы

Адгезия спор
 M. Lars. et Lomb) – место локализации экстраклеточных")
Чехлы дереворазрушающих грибов (Postia placenta (Fr.) M. Lars. et Lomb) – место локализации экстраклеточных ферментов и грибных метаболитов Предполагаемые функции чехла Гомеостаз ионов и pH, Узнавание и адгезия к субстрату, Концентрирование, запасание и транспорт продуктов деградации Защита от высыхания, Подготовка субстрата к деградации Регуляция процесса разрушения Увеличение поверхности для аэробного дыхания(Highley 1987; Eriksson 1988; Ruel et al. 1990)

Чехол у Laetiporus sulphureus

Чехол у Pleurotus pulmonarius выполняет важную роль в межклеточных контактах

Капсула дрожжей Функции капсулы дрожжей: -прикрепление клеток к поверхности твердого субстрата, -резервный источник питания, -улучшение водного снабжения клетки (в том числе защита от высыхания) -в природных местах обитания – места аккумуляции спутников и ассоциантов грибов.

Представляет собой глюкуроноксиломаннан и составляет до 88% от массы сухих веществ капсулы Капсула Cryptococcus neoformans играет главную роль в вирулентности

Грибная стенка как клиническая мишень Антигрибные препараты мишенью которых является синтез хитина ограниченно используются в случаях, даже когда хитин имеет большое значение в структуре стенки грибов и это делает его замечательной мишенью. Имеются природные ингибиторы синтеза хитина никкомицины и полиоксины, и их синтетические производные. Это аналоги субстрата хитинсинтазы, UDP-N-ацетилглюкозамин, и действует как конкурентные ингибиторы хитин синтазы. Они могут быть эффективными при использовании в сочетании с другими противогрибными агентами, но как правило неэффективны сами по себе (Bowman & Free, 2006).

Эффективной мишенью синтеза стенки грибов для коммерческих важных антигрибных агентов является β1,3-глюкан синтаза, которая ингибируется эхинокандинами. Эхинокандины вызывают к округлению/набуханию грибных клеток и лизису стенок в местах активного синтеза клеточных стенок.
20260-cell_wall_lekciya_1-2011.ppt
- Количество слайдов: 68