
КАЛЬЦИЙ-ЗАВИСИМЫЕ ПУТИ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ Роль












 В ПРИСУТСТВИИ ИОНОВ КАЛЬЦИЯ СВЯЗЫВАЮТ ФОСФОЛИПИДЫ")

 КАЛЬСЕКВЕСТРИН В поперечно-полосаты х")
















 PI-4 - фосфат ( PIP ) PI-4, 5")




")


")

4_sa_5.ppt
- Размер: 9.4 Мб
- Автор:
- Количество слайдов: 54
Описание презентации КАЛЬЦИЙ-ЗАВИСИМЫЕ ПУТИ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ Роль по слайдам
КАЛЬЦИЙ-ЗАВИСИМЫЕ ПУТИ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ
Роль ионов кальция в клетках • Выделение нейромедиаторов • Вторичный посредник Расмуссен, 1970 • Сокращение мышц
Большинство агонистов вызывает повышение внутриклеточной концентрации ионов кальция (например, ангиотензин II , эндотелин I , тромбоксан, брадикинин, гистамин, серотонин и др. ) Гораздо меньше агентов, приводящих к снижению внутриклеточной концентрации ионов кальция (например, простациклин, простагландин Е 1, Na -уретический пептид)Известно более 30 БАВ (гормоны, факторы роста, простагландины, цитокины), действие которых опосредовано ионами кальция.
Роль ионов кальция как вторичного посредника • Низкая внутриклеточная концентрация ионов кальция ( 10 -7 М) • Возможность быстро повысить внутриклеточную концентрацию (до 0, 6 – 2 мк. М) ионов кальция
БАВ Вторичные посредники: IP 3 R R Ca 2+Потенциалоза- висимые каналы Ионный обменник и насос ATP Ca 2+ Na +K + Ca 2+ s. r. Кальций-связ ывающие белки. ПУТИ ПОСТУПЛЕНИЯ И УДАЛЕНИЯ ИОНОВ КАЛЬЦИЯ Рецептор-за висимые каналы
МОЛЕКУЛЯРНАЯ СТРУКТУРА Са 2+ -КАНАЛА L — ТИПА ВЗАИМОДЕЙСТВИЕ СУБЪЕДИНИЦ КАНАЛА ДОМЕННАЯ ОРГАНИЗАЦИЯ КАНАЛА
Характеристики Ca 2+ как вторичного посредника 1. В ответ на действие БАВ активируются структуры, которые поставляют ионы кальция в клетку. 2. В ответ на БАВ увеличивается [Ca 2+ ] in. 3. Снижение активности Ca 2+ -АТФазы приводит к усилению эффекта ионов Ca 2+. 4. Эффект БАВ воспроизводится с помощью Ca 2+ -ионофора. 5. Повышение [Ca 2+ ] in предшествует биологическому эффекту.
Кальций-связывающие белки I. Кальций-связывающие белки с ферментативной активностью II. Кальций-фосфолипид-связывающие белки III. Кальций-запасающие белки IV. Кальций-связывающие белки в составе ионных каналов и др. ион-транспортных систем V. Регуляторные кальций-связывающие белки, не проявляющие ферментативной активности
кальпаин кальцинейрин Мультисубстратная протеаза Протеинфосфатаза I. Кальций-связывающие белки с ферментативной активностью
КАЛЬПАИН Гетеродимер: большая каталитическая и малая регуляторная субъединицы. гидролизует белки цитоскелета, ядерные белки, мембранные рецепторы, многие ферменты, транспортные белки
КАЛЬЦИНЕЙРИН Дефосфорилирует различные функционально важные фосфобелки по серину и треонину Гетеродимер, состоит из регуляторной и каталитической субъединиц, которые, в свою очередь, имеют несколько доменов. Широко представлена в мозге
II. Кальций-фосфолипид-связывающие белки АННЕКСИНЫ (известно более 10) В ПРИСУТСТВИИ ИОНОВ КАЛЬЦИЯ СВЯЗЫВАЮТ ФОСФОЛИПИДЫ МЕМБРАН, ТЕМ САМЫМ • УЧАСТВУЮТ В ПРОЦЕССАХ СЛИЯНИЯ И АГРЕГАЦИИ МЕМБРАН • ИНГИБИРУЮТ ФОСФОЛИПАЗУ А 2 • КОНТРОЛИРУЮТ ПРОЛИФЕРАЦИЮ И ДИФФЕРЕНЦИРОВКУ КЛЕТОК Содержат 4 Са 2+ -связывающих домена
Во время апоптоза клетки высвобождают фосфатидилсерин на клеточной поверхности. Аннексин V , являющийся фосфолипид-связывающим протеином, в присутствии ионов кальция селективно, с высокой аффинностью, связывает фосфатидилсерин. Он проявляет очень низкую аффинность к таким фракциям фосфолипидов, как фосфатидилэтаноламин, сфингомиелин и фосфатидилхолин. Такой профиль связывания позволяет использовать аннексин V в качестве высокоспецифичного агента для определения апоптических клеток.
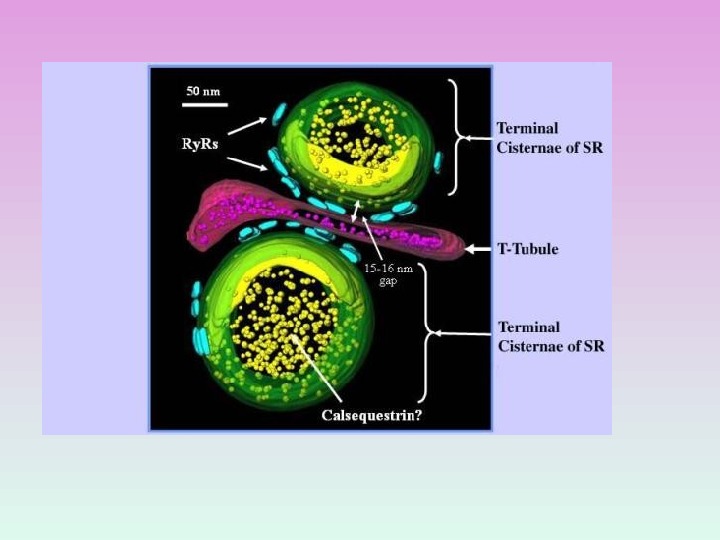
III. Кальций-запасающие белки СИСТЕМА ПАССИВНОГО ЗАБУФЕРИВАНИЯ ИОНОВ КАЛЬЦИЯ (примеры) КАЛЬСЕКВЕСТРИН В поперечно-полосаты х мышцах КАЛЬРЕТИКУЛИН В СПР
КАЛЬСЕКВЕСТРИН
IV. Кальций-связывающие белки в составе ион-транспортных систем Са 2+ зависимые калиевые каналы Са 2+ зависимые хлорные каналы Na + /Ca 2+ обменник Са 2+ -АТФаза Са 2+
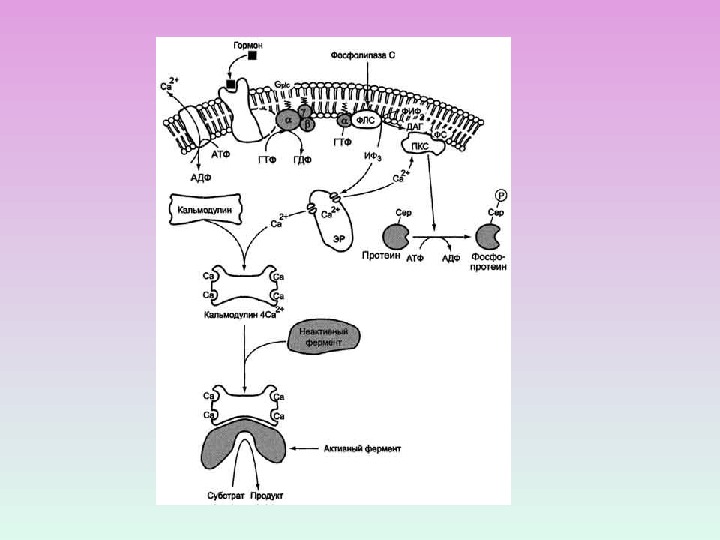
V. Регуляторные кальций-связывающие белки, не проявляющие ферментативной активности Впервые обнаружен Ченгом в мозге КРС в качестве Са 2+ -зависимого активатора фосфодиэстеразы циклических нуклеотидов. Впоследствии обнаружен во всех эукариотических клетках. Центральное место среди этих белков занимает КАЛЬМОДУЛИН
Является “ E – F hand ”- протеином. В структуре белка было обнаружено 6 -спиралей ( A, B, C, D, E, F ). В петлях полипептидной цепи между участками C и D , E и F находятся функциональные группы с высоким сродством к кальцию. Спирали E и F расположены как вытянутые указательный и большой пальцы- ион Са 2+
E — F hand белки разделяются на группы в зависимости от количества Са-связывающих центров. 4 Са-связывающих центра, кроме кальмодулина, имеют тропонин С (из скелетных мышц), легкие цепи миозина, кальцинейрин
Ион кальция
Связывающие Са 2+ участки содержат остатки кислых аминокислот и обогащены атомами кислорода. 6 – 8 атомов кислорода образуют ячейку, соответствующую размеру иону кальция ( r 0, 099 нм)
Са 2+ -кальмодулин-зависимые Са-АТФаза ФДЭ Диссоциация микротрубочек. Сокращение ГМК Са-Са. М-зависимая протеинкиназа Киназа легких цепей миозина (в ГМК) Аденилатциклаза Клеточная пролиферация ферменты процессы Высвобождение нейромедиаторов
КОМПЛЕКС КАЛЬЦИЙ-КАЛЬМОДУЛИН ПРЯМОЕ ВЗАИМОДЕЙСТВИЕ С ФЕРМЕНТОМ-МИШЕНЬЮ ЧЕРЕЗ Са 2+ -КАЛЬМОДУЛИН-ЗАВИСИМУ Ю ПРОТЕИНКИНАЗУ
ПРЯМОЕ ВЗАИМОДЕЙСТВИЕ С ФЕРМЕНТОМ-МИШЕНЬЮ
ПРЯМОЕ ВЗАИМОДЕЙСТВИЕ КОМПЛЕКСА Са 2+ – КАЛЬМОДУЛИН С ФЕРМЕНТОМ-МИШЕНЬЮ
АКТИВАЦИЯ ФЕРМЕНТОВ – МИШЕНЕЙ ЧЕРЕЗ Са 2+ -КАЛЬМОДУЛИН-ЗАВИСИМУЮ ПРОТЕИНКИНАЗУ
С а Са 2+ канал Са 2+ -КАЛЬМОДУЛИН- ЗАВИСИМАЯ ПРОТЕИНКИНАЗА
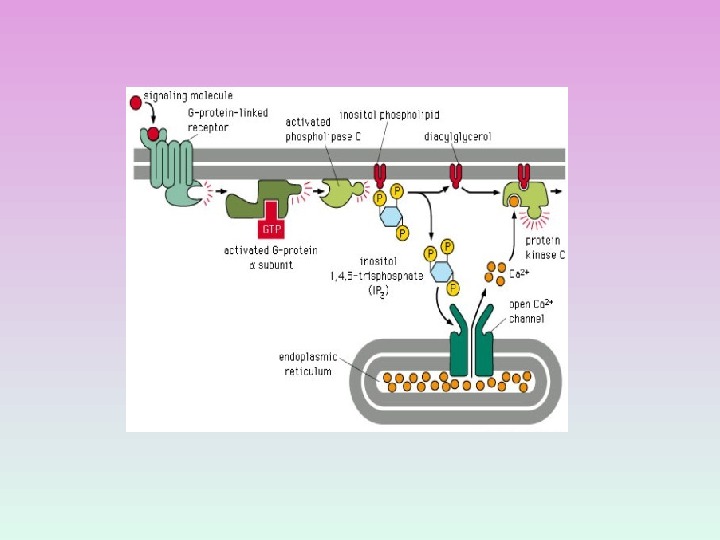
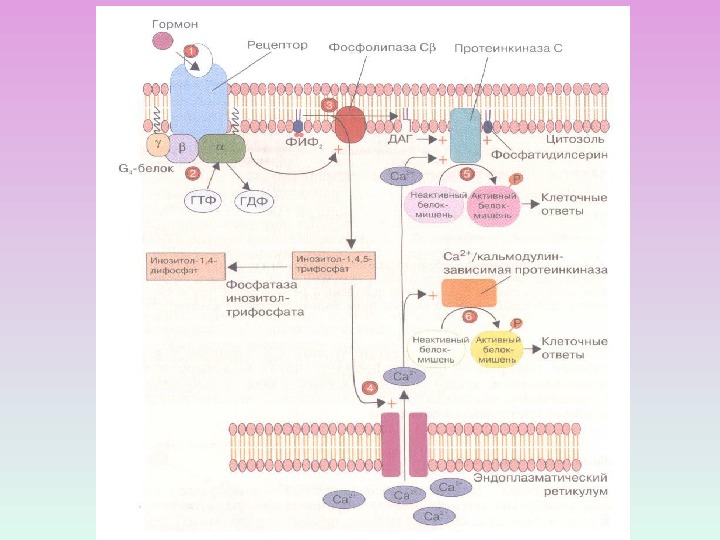
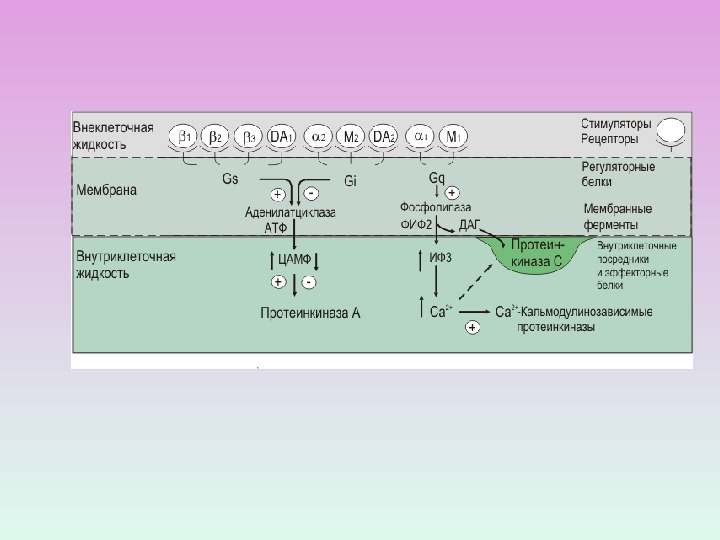
СИГНАЛЬНАЯ СИСТЕМА, ОПОСРЕДОВАННАЯ ИОНАМИ КАЛЬЦИЯ И ПРОДУКТАМИ ГИДРОЛИЗА МЕМБРАННЫХ ФОСФОИНОЗИТИДОВ
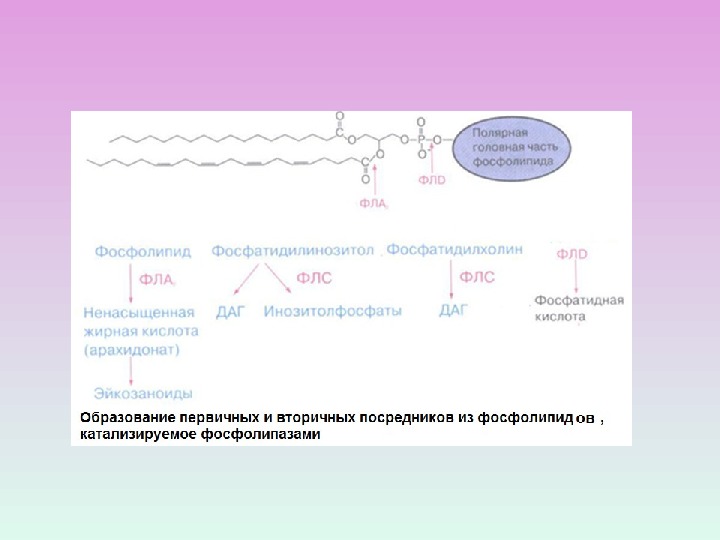
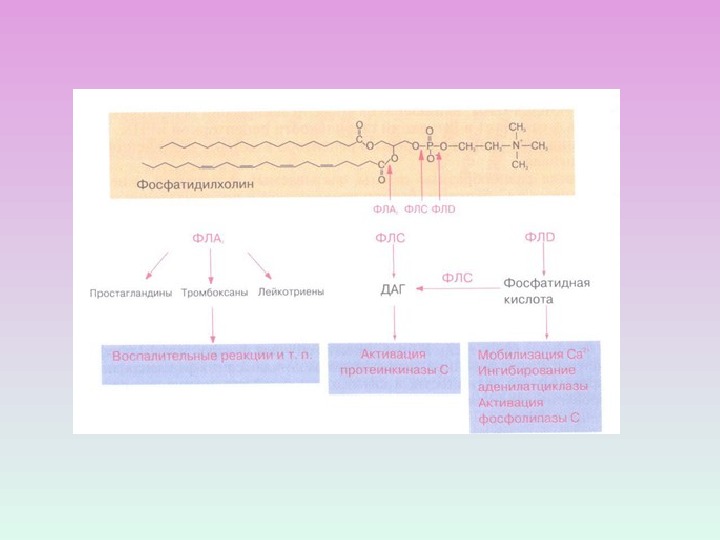
PL А 1 – гидролиз эфирной связи в положении 1 ( отщепляет SN-1 ацильную цепь). PL А 2 – гидролиз эфирной связи в положении 2 ( отщепляет SN-2 ацильную цепь, образование свободных жирных кислот)PL С – гидролиз эфирной связи в положении 3 ( гидролизует связь между глицериновым остатком фосфолипида и полярной фосфатной группой ) PL D — гидролизует связь между фосфатной группой и спиртовой группой ВИДЫ ФОСФОЛИПАЗ
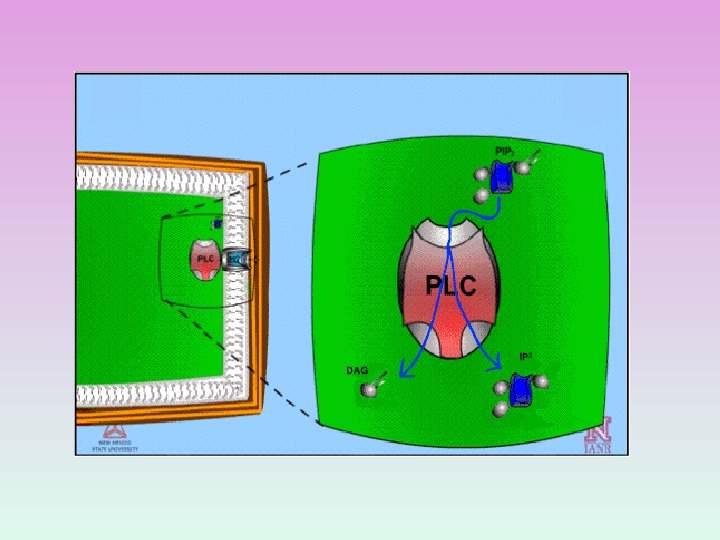
ФОСФОЛИПАЗА С ГИДРОЛИЗУЕТ МИНОРНЫЕ ФОСФОЛИПИДЫ МЕМБРАН — ФОСФОИНОЗИТИДЫ
МЕТАБОЛИЗМ МЕМБРАННЫХ ФОСФОИНОЗИТИДОВ Фосфатидилинозитол( PI ) PI-4 — фосфат ( PIP ) PI-4, 5 — бифосфат ( PIP 2 )
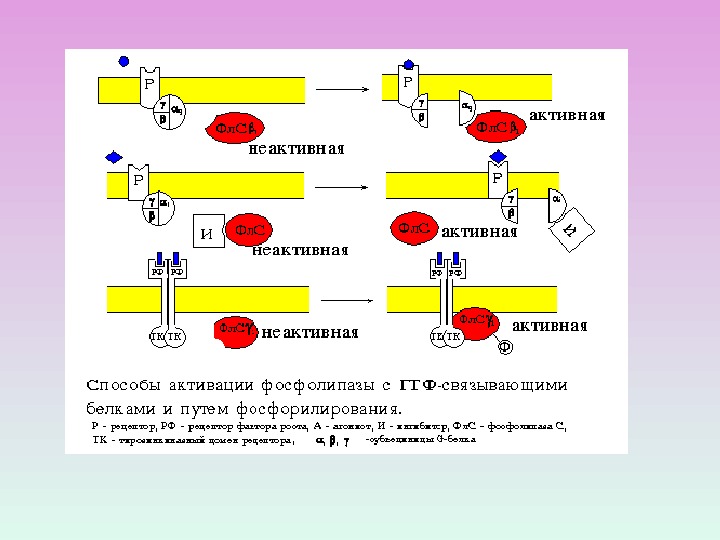
ИЗОФОРМЫ ФОСФОЛИПАЗЫ С
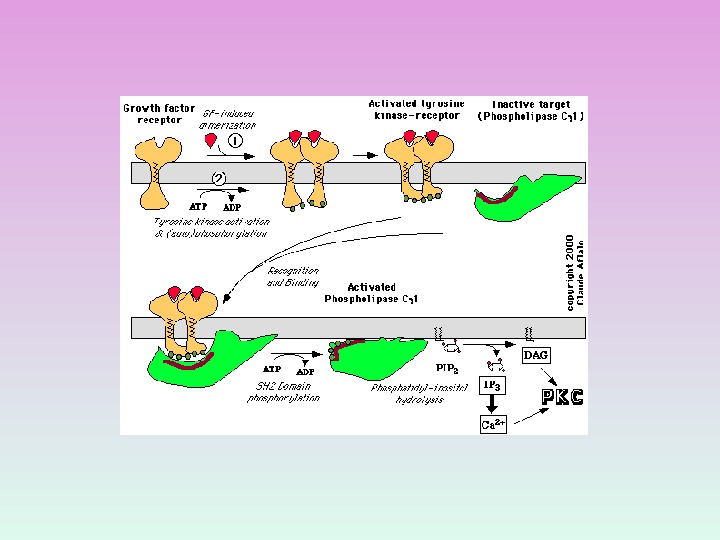
ФОСФОЛИПАЗА С: изоформы и ФОСФОЛИПАЗА С активируется через рецепторы, связанные с G -белками ( G q белок) ФОСФОЛИПАЗА С активируется благодаря фосфорилированию ее тирозинкиназой.
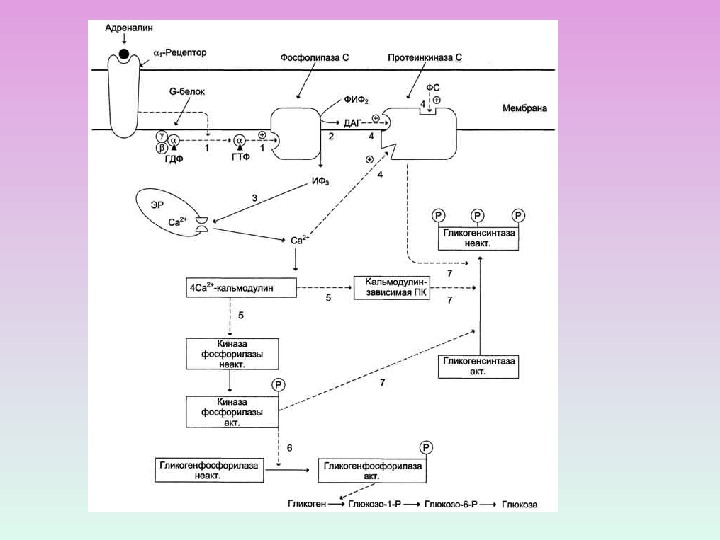
1 — адренорецепторы , мускариновые рецепторы ФОСФОЛИПАЗА С
Факторы роста ФОСФОЛИПАЗА С
PI-4, 5 — биф осфат ( PIP 2 )
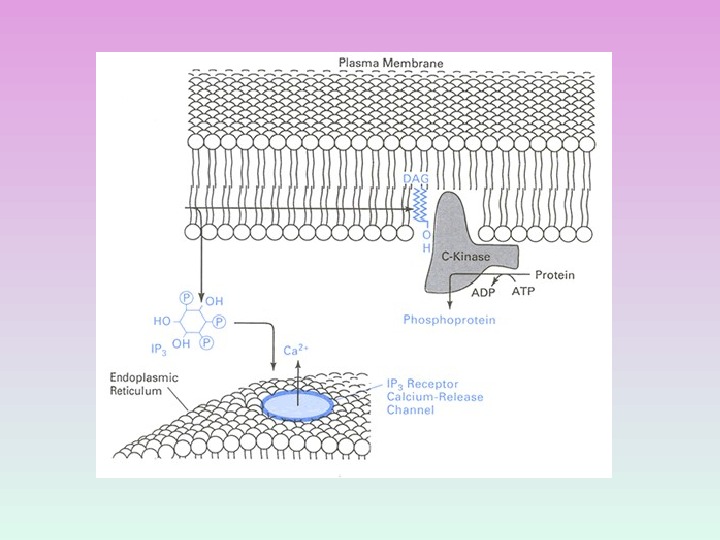
ПРОДУКТЫ ГИДРОЛИЗА ФОСФОИНОЗИТИДОВ: ВТОРИЧНЫЕ ПОСРЕДНИКИ Диацилглицерол – гидрофобная молекула, которая остается в мембране. Под действием фосфолипазы А 2 дает арахидоновую кислоту, которая является предшественником эйкозаноидов. Инозитолтрифосфат – гидрофильная молекула, которая диффундирует в цитозоль и взаимодействует с рецепторами ретикулума
ПРОЦЕССЫ, В КОТОРЫХ УЧАСТВУЕТ ПРОТЕИНКИНАЗА С Секреция Сокращения ГМК Агрегация тромбоцитов Фосфорилирование рецепторов
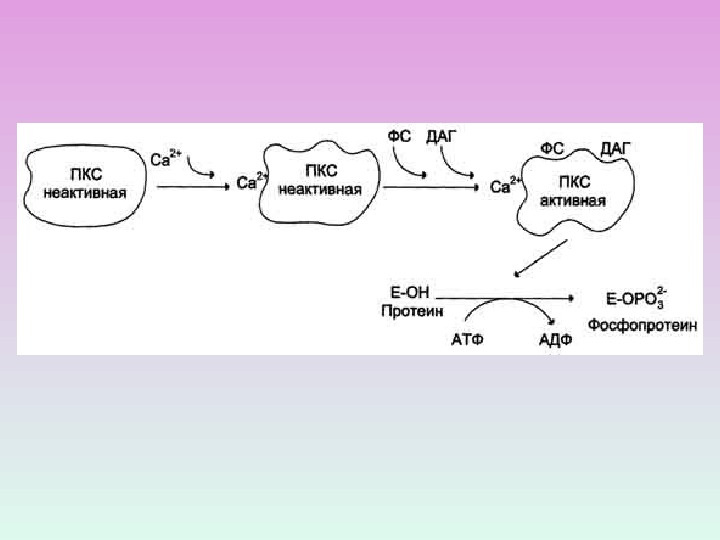
R С 2 Zn 2+ Протеинкиназа С состоит из двух доменов – регуляторного (R) и каталитического (C) Регуляторный домен: мотив цинковые пальцы и участок с высоким сродством к Са 2+ДАГ Са 2+ Каталитический домен: центр, связывающий АТФ и белок-субстрат. АТФ Белок-су бстрат Неактивная протеинкиназа С – цитозольная , при активации становится мембранносвязанной